|
|
|
|
Celule prezentatoare de antigen (APC)
=celule ce joaca un rol esential in initierea in organele limfoide secundare (ggl limfatici, splina) a unor raspunsuri imune, fie umorale, fie celulalre fata de Ag timodependente.
Ag timodependente
participa doar in initierea raspunsurilor imune
raspunsurile imune se desfasoara doar in organele limfoide secundare
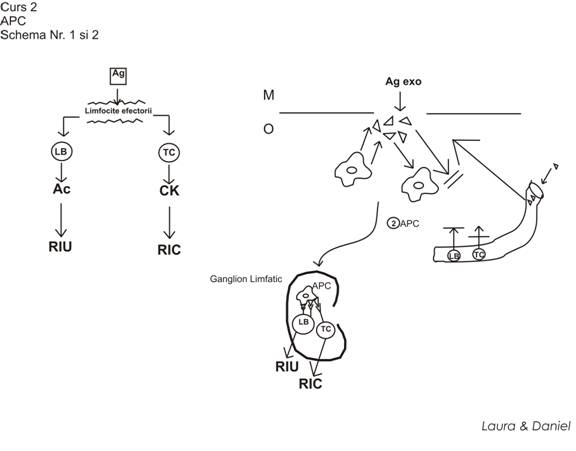
conditia de baza in declansarea raspunsurilor imune consta in stimularea de catre Ag inductor a limfocitelor efectorii
limfocite efectorii: Limfocite B, Limfocite Tc
activarea limfocitelor efectorii NU se realizeaza niciodata per primam, ci activarea lor se realizeaza prin parcurgerea a mai multe etape => raspunsurile imune ce apar sunt procese multifazice
Etapele de principiu ale raspunsurilor imune fata de Ag timodependente
ele se pot acumula in diverse tesuturi
ele pot proveni din sursa externa (Ag exogene) daca strabat barierele dintre mediu si organism (t. cutanat, epiteliu digestiv, epiteliu respirator)
sau din sursa endogena (eliberate ca urmare a generarii unor procese autoimune, Ag tumorale care se pot acumula ca urmare a eliberarii lor de catre celulele tumorale - Ag eliberate la distanta foarte mare de tesutul tinta, ele sunt lansate in circulatie si ulterior pot difuza extravascular cu acumulare in tesuturi si organe)
limfocitele efectoare pot fi localizate: - in circulatie
- in organele limfoide, unde sunt prezente marea majoritate a clonelor de LB si LT
Ag NU poate activa limfocitele circulante
LB circulante NU pot fi activate pentru ca obisnuit ele NU difuzeaza extravascular
LTc pot trece extravascular, dar NUMAI in conditii speciale (daca ele sunt celule cu memorie imunologica)
Ag pot activa limfocitele din organele limfoide (ggl, splina). Ele ajung acolo ca urmare a faptului ca sunt preluate si transportate acolo de APC
-APC se localizeaza in organele limfoide si aici incep sa prezinte pe membrana lor Ag transportat pana la acest nivel
- LTc => RIC
APC
in functie de profilul lor functional se impart in doua categorii:
celule produse de maduva hematogena, ce lanseaza in circulatia sistemica elemente tinere sub forma de monocite
acestea, dupa un foarte scurt tranzit intracirculator (24-36h), trec in diverse tesuturi si organe, fiind atrase sub actiunea unor factori chemotactici
in final => macrofage care devin macrofage activate
ajunse in stadiul de macrofag activat pot indeplini functia de APC
functia de APC trece obligatoriu prin 5 etape:
1) Captarea Ag declansatoare
macrofagele capteaza de regula NUMAI Ag corpusculate (bacterii, virusuri, mici paraziti). NU capteza Ag solubile. Macrofagele capteaza Ag corpusculare aflate in 2 stari: opsonizate (mai frecvent) si neopsonizate.
captarea e un proces mediat receptorial
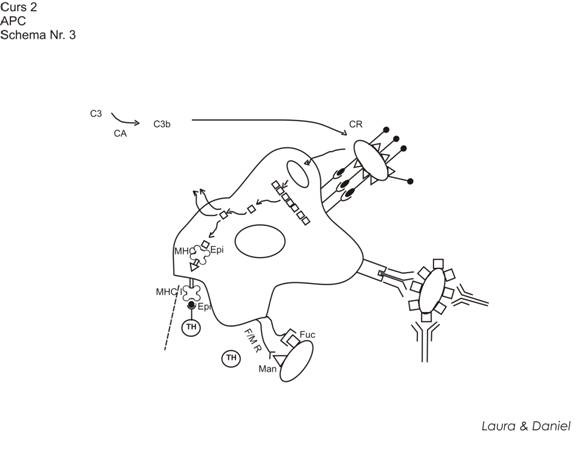
Opsoninele factori imunologici solubili care invelesc sau tapeteaza Ag corpusculate si favorizeaza captarea Ag de APC
Exista 2 opsonine de importanta: C3b a complementului (fragment rezultat din activarea factorului C3 al complementului). Ea e pusa in libertate in urma functionarii continue a caii alterne a complementului. C3b are ca particularitate faptul ca se ataseaza de orice membrana NON-SELF => depunerea ei pe suprafata bacteriei. Macrofagele capteaza foarte usor aceste bacterii pentru ca dispun pe suprafata lor de receptori ce pot angaja legaturi cu componenta C3b a complementului (CR)
IgG se depun pe suprafata bacteriilor ca urmare a recunoasterii determinantului Ag de suprafata. IgG angajeaza legaturi cu ei prin intermediul capatului Fab al IgG. Astfel IgG proemina in lichidul interstitial prin capatul Fc. Macrofagele pot angaja foarte usor aceste legaturi prin Fc al IgG (au receptori pt Fc al IgG = receptori FcγR)
Mai putin eficient pot fi captate si Ag neopsonizate ca urmare a faptului ca macrofagele au recptori pe suprafata capabili sa recunoasca unele oligozaharide prezente in peretele Ag (fucozil, manozil). Acesti receptori se numesc F/M-R.
Cei mai importanti receptori sunt CR si FcγR (receptori opsoninici)
captarea e un proces imunologic specific. Macrofagele, prin receptorii de suprafata NU vin niciodata in contact direct cu determinantii Ag de suprafata ai bacteriilor => proces imunologic specific.
2) Endocitarea Ag captate
se realizeaza ca urmare a emiterii de pseudopode si constituirea unor vacuole
e un proces inalt consumator de energie
3) Prelucrare/procesare Ag endocitate, mai ales in compartimentul lizozomal
aici se realizeza fragmentarea Ag nativ, scindarea lui cu generare a numeroase fragmente ce poara denumirea de epitopi
din mai multi epitopi, macrofagele realizeaza selectia epitopilor imunodominanti (cei mai capabili de declansarea unui raspuns imun)
4) Conserverea unei mici parti din epitopii imunodominanti in anumite subsectoare ale macrofagului
ea e importanta pentru ca din cantitatea conservata ulterior macrofagul va elibera mici cantitati de epitopi, ceea ce conduce la intretinerea in timp a raspunsului imun
5) Cuplarea majoritatii epitopilor imunodominanti in moleculele MHC => in final complexe MHC-epitopi si prezentarea Ag
complexele MHC-epitopi sunt prezentate pe membrana macrofagelor
prezentarea se realizeza in cuplu cu moleculele MHC I, MHC II
epitopii sunt prezentati catre Limfocitele TH. Ele cunosc doar epitopii care sunt prezentati in complex cu MHC II
din toatlul macrofagelor:
periferice - 75% pot sintetiza MHC II
splenice - 50% pot sintetiza MHC II
hepatice - 25% pot sintetiza MHC II
daca macrofagele elaboreaza si MHC I si MHC II => ele sunt implicate in generarea unor raspunsuri imune
daca ele produc doar MHC I NU sunt implicate in generarea de raspunsuri imune, ele participa in reactii inflamatorii
cele mai importante etape pentru functia de APC sunt prima si a cincea
functia de APC e conditionata de receptorii de suprafata si de producerea de MHC II
Functia de ansamblu a macrofagelor
transportul Ag tisulare prin limfaticele aferente catre ariile timodependente (populate de Limfocitele TH) ale organelor limfoide secundare. Aceste arii sunt reprezentate de zona paracorticala in ggl limfatici si de PALS intern in splina (PALS = stratul limfoid periarterial intern)
celule produse de maduva hematogena ce au ca principala caracteristica morfologica faptul ca in conditii de repaus prezinta prelungiri citoplasmatice sau pseudopode
pseudopodele au 3 caracteristici:
o sunt numeroase sau foarte numeroase
o sunt fine, efilate
o sunt lungi, depasesc de 4 - 5 ori diametrul celulei propriu-zise
seamana morfologic cu celulele dendritice din SNC
indeplinesc functie de APC pentru ca:
o in general dispun de receptori de captare (CR, FcγR)
o in general ele elaboreaza molecule MHC
ele NU reprezinta o clasa omogena, ci una foarte heterogena atat morfologic, cat si functional
se disting 6 clase distincte, impartite in 3 clase mai mari (vezi tabel)
Nelimfoide
1. CD Langerhans
distribuite in tesutul cutanat (limita demo - epidermica) unde formeaza un strat celular aproape continuu; submucoasa bucala; submucoasa tractului digestiv; submucoasa si mucoasa colului uterin
aspect atipic dendrititc. Ele sunt celule stelate (prezinta pseudopode putine si groase)
ele dispun de receptori CR, FcγR din abundenta, insa NU elaboreaza MHC II
functia lor: transporta Ag de la nivelul t. cutanat sau alte locatii catre ariile timodependente ale organelor limfoide secundare. Prezinta Ag catre LTH
2. CD Interstitiale
ele sunt distribuite ubicuitar la nivelul interstitiilor celulare
au aspect morfologic atipic dendritic (sunt stelate)
seamana cu CD Langerhans. Prezinta receptori opsoninici, dar NU elaboreaza MHC II
ele transporta Ag tisulare catre ariile timodependente ale organelor limfoide secundare
Limfoide
3. CD Interdigitate
sunt distribuite NUMAI in ariile timodependente ale organelor limfoide secundare
sunt celule tipic dendritice
ele sunt cele mai mari producatoare de MHC II, insa NU au receptori de captare Ag (CR, FcγR)
4. CD Foliculare
pentru ca sunt prezente in foliculii limfoizi (aglomerari de limfocite B). Ele sunt in zone bursodependente: cortexul ganglionar, respectiv PALS extern a splinei
expun pe suprafata lor in cantitati enorme receptori de captare (CR, FcγR). NU elaboreaza MHC II
joaca un rol important in concentrarea in zone bursodependente a Ag declansatoare, ceea ce favorizeaza recunoasterea Ag de Limfocitele B. Ele au o functie de filtru celular
Circulante
5. CD Sangvine
prezente NUMAI in circulatie, dar in numar mic (sub 0,1% din toatlul PMN)
NU dispun de nici una din categoriile de molecule prezente pe suprafata
6. CD Valuroase
celule aplatizate care prezinta pe flancuri oscilatii sau ondulatii ale membranei
sunt prezente NUMAI in limfa
NU dispun de nici un fel de receptori si nici NU elaboreaza molecule de MHC II
|
|
CD |
CR |
FcγR |
MCH II |
|
Nelimfoide (intratisulare) |
CD Langerhans |
+++ |
+++ |
― |
|
CD Interstitiale |
+++ |
+++ |
― |
|
|
Limfoide |
CD Interdigitate |
― |
― |
+++++ |
|
CD Foliculare |
+++++ |
+++++ |
― |
|
|
Circulante |
CD Sangvine |
― |
― |
― |
|
CD Valuroase |
― |
― |
― |
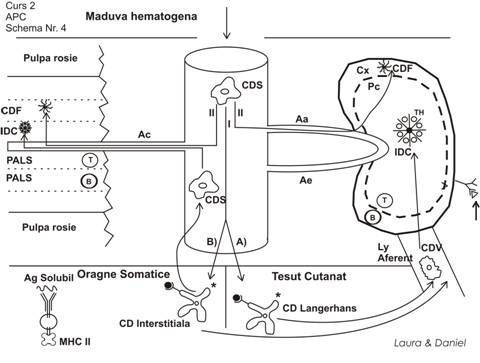
Se considera ca aceste 6 tipuri de celule NU reprezinta entitati morfologice in sine, ci toate provin dintr-o singura celula progenitoare, care in functie de ariile pe care le traverseaza, isi modifica aspectul morfologic, de aceea discutam despre traficul si filiatia celulei dendritice.
Maduva osoasa lanseaza celule dendritice in circulatie, apoi ele trec in tesuturi si organe.
De asemenea din circulatie trec in ggl locoregionali. Se considera ca parenchimul ggl prezinta la exterior o zona - cortex - bursodependenta, iar la interior o zona - paracortex - timodependenta.
In splina ajung prin arteriola centrala. In jurul ei se afla pulpa alba. Ea e populata de leucocite. In jurul pulpei albe se afla pulpa rosie. Pulpa alba e alcatuita din 3 masoane concentrice:
1. PALS intern (timodependenta)
2. PALS extern (bursodependenta)
3. Zona marginala
Initial sunt produse CDS, care sunt ovalare. Ele pot urma 2 traficuri:
1. traficul intratisular
a. tesut cutanat
aici isi modifica aspectul morfologic (devin stelate) = CD Langerhans
ele prezinta pe suprafata receptori de captare Ag (CR, FcγR) prin care pot capta diverse Ag
apoi se activeaza si incep sa migreze, transportand Ag
ele iau calea limfaticelor aferente catre ggl locoregionali
in limfa ele isi modifica aspectul in CD Valuroase si patrund in ggl locoregionali, unde se stabilesc NUMAI in paracortex
aici isi modifica aspectul in celule tipic dendritice (CD Interdigitate). Ele produc MHC II prin care prezinta Ag catre TH, care se inclaveaza printre prlungirile lor => activare TH cu declansarea raspunsului imun
b. organe somatice
aici isi modifica aspectul si devin stelate (CD Interstitiale)
functie de captare Ag (prezinta CR, FcγR)
ele se activeaza si pot urma 2 tipuri de traficuri:
calea limfaticelor aferente se transforma in CD Valuroase si sub aceasta forma Ag e transportat in ggl locoregionali, unde ele se transforma in CD Interdigitate
calea antidromica si reintra in circulatie, se transforma din nou in CD Sangvine. Sub aceasta forma patrund in splina, luand calea arteriolei centrale si se fixeaza in PALS intern, unde se transforma morfologic in CD Interdigitate si prezinta Ag catre LTH
2. traficul limfoid
CD Sangvine pot patrunde direct in organele limfoide
in ggl, unde se opresc in cortex, luand aspectul de CD Foliculare
in splina, prin arteriola centrala si se fixeaza in PALS extern, unde devin CD Foliculare
pot functiona ca APC pentru ca prezinta pe suprafata lor receptori de captare Ag (CR, FcγR si BCR)
BCR capteaza Ag solubile
In anumite etape LB pot elabora si expune molecule MHC II
Moleculele prezentatoare de Ag (MHC I si MHC II)
1. MHC I
distribuite ubicuitar, pe membrana tuturor celulelor, cu exceptia eritrocitelor si a trombocitelor
sunt heterodimeri, formati din 2 lanturi diferite, dar asociate intre ele, orientate cu capetele N-terminale in afara celulei si cele C-terminale in interior. Ele se noteaza cu α si β.
α e un lant mare, cu GM de aproximativ 40-45 kda,
β are GM de aproximativ 12 kda (β2 microglobulina)
o lantul α
foarte lung, NU este liniar
el e constituit din 3 bucle (domenii), taiate prin punti sulfurice intracatenare
buclele sunt formate din 100-110 aa
cele 3 domenii se noteaza cu α1, α2, α3
α1α2 sunt constituite din secvente de aa variabile sau extrem de variabile (domenii polimorfe). Ele sunt pozitionate fata in fata si constituie o cavitate de dimensiuni mici = situs de prezentare a Ag
α3 au o structura constanta pe toate celulele unui individ. E considerat un Ag self. La acest lant se asociaza lantul β
o lantul β
pozitionat extracelular
realizeaza legaturi covalente cu α3
are structura constanta la toti indivizii aceleiasi specii
Functiile MHC I:
prezentarea Ag endogene
responsabile prin α3 de declansarea reactiilor de respingere a grefelor de tesuturi si organe
cooperarea dintre APC, pe suprafata carora se poate afla, si LTc sau LTS. In timpul cooperarii dintre ele se angajeaza legaturi prin MHC I cu un contraligand prezent pe membrana LTc si LTS (el se numeste CD8). Angajeaza legaturi cu domeniul α3.
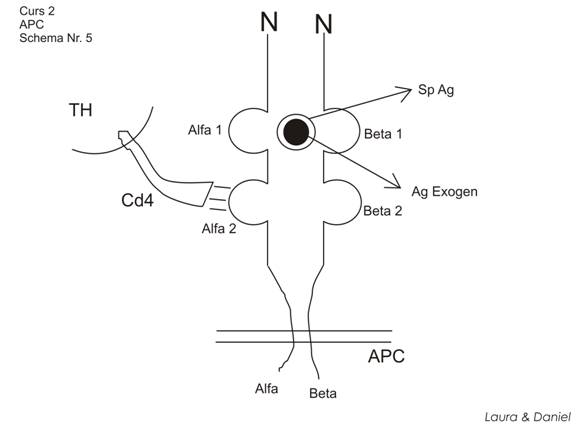
2. MHC II
distribuite restrans (pe membranele celulelor participante la raspunsurile imune: APC, LB sau LT)
sunt heterodimeri, formati din 2 lanturi cu GM apropiata
α - GM 32-34 kda
β - GM 27-28 kda
lanturile sunt organizate in bucle sau domenii. Pe fiecare lant exista 2 domenii (α1, α2, β1, β2), dinspre capatul N-terminal spre cel C-terminal.
α1, β1 au structura variabila sau hipervariabila (domenii polimorfe). Ele sunt pozitionate fata in fata, se onduleaza => o cavitate = situs prezentator de Ag
α2, β2 sunt domenii monomorfe
 Functiile
MHC II:
Functiile
MHC II:
prezentarea Ag exogene (provenite prin fagocitare)
pot avea o contributie la declansarea unor reactii de respingere a grefelor (α2)
cooperare APC si LTH => legaturi intre MHC II si un contraligand prezent tipic pe TH (CD4) => legaturi CD4- α2
Mecanismele prezentarii selective a Ag de catre moleculele MHC
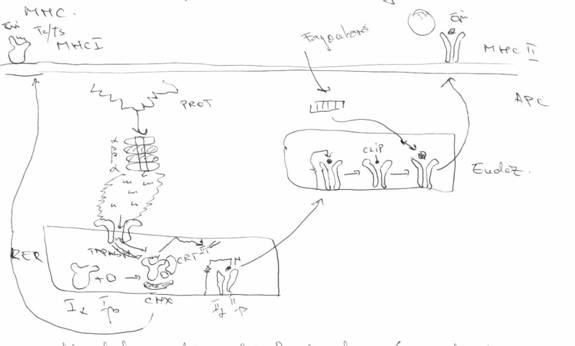
1. MHC I
prezinta Ag endogene (provenite din citoplasma APC). La origine ele sunt proteine citoplasmatice ce pot fi autoantigene, Ag tumorale, Ag rezulate din metabolizarea bacteriilor cu habitat intracelular, din diverse categorii de virusuri ce colonizeaza citoplasma
proteinele trebuiesc prelucrate => un compartiment enzimatic (proteozomi = materiale enzimatice non-lizozomale, constituite din unitati sub forma de inel - 4 - α la exterior si β la interior). α au rol imunoreglator, iar β sunt unitati enzimatice ce contin diverse tipuri de endopeptidaze
mici fragmente peptidice = epitopi. Ei vor fi tranlati din citoplasma in RER, unde se elaboreaza moleculele MHC I
translarea NU se face prin difuziune, ci prin sisteme transportoare ce se numesc TAP1/2 (transportorul asociat prezentarii Ag)
proteozomii sunt ancorati la TAP prin intermediul unor proteine de legatura LMP (low mass proteins) si sunt 2 varietati: LMP-2 (2 kda) si LMP-7 (7 kda)
epitopii sunt incarcati pe MHC I, care sunt eliberati initial ca lanturi individuale (Iα si Iβ). Ele se asociaza => molecule MHC I (dar lanturile NU au afinitate mare intre ele). Ele sunt solidarizate ca urmare a asocierii unor chaperine (CALNEXINA). Inca NU e capabil sa fixeze epitopii. Cea de-a doua chaperina ii da capacitatea aceasta. Ea induce modificari conformationale in α, soldate prin deschiderea larga a situsului pentru prezentarea Ag (CALRETICULINA) - CR5 permite atasarea complexului la sistemul TAP (creste afinitatea Iα pentru tapsina, care leaga MHC I la sistemul TAP) => epitopii sunt incarcati si complexul MHC I e adus la suprafata, iar epitopii vor fi recunoscuti de LTc sau LTS.
2. MHC II
2 lanturi IIα si Iiβ, aflate in RER. La ele se asociaza lantul IIi(inhibitor). Prin capatul N-terminal el obtureaza situsul pentru prezentarea Ag al acestor molecule => epitopii NU pot fi incarcati pe MHC II. Cand se cupleaza sunt translate in endozomi.
in endozomi: lantul IIi: se indeparteaza capatul C-terminal, el ramanand atasat prin capatul N terminal (CLIP - peptidul inhibitor divizat prin clivaj)
lantul CLIP e indepartat => situsul prezentator de Ag devine liber => moleculele pot fixa epitopii rezultati prin calea endocitara (fagocitoza)
la incarcare moleculele sunt aduse la suprafata si epitopii prezentati vor fi recunoscuti doar de LTH