|
|
|
|
ORGANIZAREA MATERIALULUI GENETIC LA BACTERII
Genomul bacterian este alcatuit din repliconi, care sunt formatiuni genetice ce se pot replica independent. Astfel, se descriu:
. cromozomul bacterian
. elemente genetice extracromozomiale - plasmide
- genomul bacteriofagilor
. elementele genetice transpozabile - fragmentele de insertie (IS)
- transpozonii (Tn).
1. Cromozomul bacterian
Majoritatea genelor bacteriene se gasesc intr-un cromozom haploid, echivalent nuclear care codifica informatiile absolut necesare supravietuirii speciei in conditii normale.
Cromozomul la Escherichia coli este format dintr-o molecula circulara dublu spiralata de ADN ce reprezinta 80% din greutatea lui, dintr-o componenta proteica, ARN-polimeraza care reprezinta 10% iar restul de 10% din ARNm si ARNr in curs de sintetizare.
Pe langa dispozitia dublu spiralata spre dreapta a catenelor de ADN (o spira = 10 baze), acestea sufera o suprahelicare spre stanga (o spira suprahelicata = 15 spire). Acest mod de organizare denumit "supercoil" sau "supertwist" asigura pe de o parte "impachetarea" economica a ADN si pe de alta parte o configuratie optima activitatii functinale a ADN (replicare, transcriptie, recombinare). Modelul de impachetare a nucleului bacterian a fost descris de Pettijohn si Hecht (1973) pentru E. coli si se pare ca are un caracter general la bacterii.
Formatiunile genetice extracromozomiale
1. Plasmidele
Figura 21 - Structura plasmidului
 Plasmidele sunt formatiuni
genetice autonome, extracromozomiale, libere in citoplasma, reprezentate
de molecule circulare de ADN care se replica independent de cromozom.
Denumirea de plasmide le-a fost data de Lederberg in 1952, iar
importanta lor practica a fost sesizata in 1963 de Watanabe, o
data cu descoperirea posibilitatii transmiterii prin plasmide a
rezistentei la antibiotice intre bacterii .
Plasmidele sunt formatiuni
genetice autonome, extracromozomiale, libere in citoplasma, reprezentate
de molecule circulare de ADN care se replica independent de cromozom.
Denumirea de plasmide le-a fost data de Lederberg in 1952, iar
importanta lor practica a fost sesizata in 1963 de Watanabe, o
data cu descoperirea posibilitatii transmiterii prin plasmide a
rezistentei la antibiotice intre bacterii .
Bacteriile contin foarte frecvent plasmide, descriindu-se peste 1.000 de tipuri diferite. Lungimea acestor molecule variaza intre 1-150 mm si greutatea moleculara intre 2-3 milioane (3x103 pana la 450x103 perechi de baze). Plasmidele mici se gasesc de obicei intr-o celula bacteriana in 10-40 de copii, pe cand cele mari in numar mai redus de 1-3 copii. In aceeasi celula nu pot coexista mai multe plasmide inrudite (cu mecanism comun de control al replicarii), ele fiind incompatibile, spre deosebire de cele neinrudite (cu mecanism distinct de reglare al replicarii), care sunt compatibile. Compatibilitatea exprima competitia plasmidelor pentru un anumit situs de legare in celula bacteriana.
Plasmidele pot fi:
. conjugative, care se pot transfera singure la alte bacterii, ca de exemplu plasmidele de rezistenta la antibiotice R,
. neconjugative, care nu pot parasi ele insasi bacteria de origine, ci numai prin intermediul unui alt plasmid conjugativ sau a unui bacteriofag (de exemplu plasmidul care codifica secretia de beta-lactamaza la S.aureus)
. episomi, care se pot integra prin recombinare in cromozomul bacterian, pierzandu-si astfel autonomia de replicare. Un exemplu de episom este factorul de sex F (plasmidul F sau factorul de fertilitate).
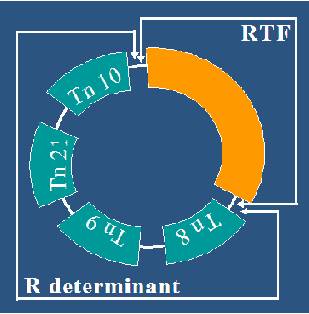
Determinantii genetici esentiali ai plasmidelor codifica informatiile legate de replicarea lor autonoma, iar cei accesorii caractere fenotipice neesentiale supravietuirii celulei bacteriene in conditii naturale: gene de transfer (tra), de rezistenta la antibiotice (factorul R), de secretie a colicinelor (factorul col), de secretie a unor toxine, de metabolizarea unor substraturi, de rezistenta la unii ioni si compusi organometalici etc. Unele plasmide nu au un efect observabil asupra fenotipului bacteriei si se numesc plasmide criptice.
Pentru medicina importanta este cunoasterea plasmidelor de virulenta si a celor care confera bacteriilor rezistenta la antibiotice.
Plasmidele de virulenta poarta determinantii genetici ai unor factori de virulenta la bacterii, ca de exemplu secretia de enterotoxina (termolabila si termostabila) si factorul de colonizare la Escherichia coli, hemolizina la Staphylococcus aureus, Streptococcus faecalis si E.coli, exfoliantina la S. aureus, gena de invazivitate la Shigella etc.
Plasmidele R -de rezistenta la chimioterapice (Factorul R) sunt molecule circulare de ADN care constau in esenta din doua regiuni genetice distincte: genele care codifica rezistenta la antibiotice "R", care pot fi unice sau multiple si genele care confera plasmidului capacitatea de a se transfera "FTR".
Rezistenta la peste 90% din tulpinile de spital este de natura plasmidica. Existenta acestor plasmide se explica prin aglomerarea mai multor gene de rezistenta R, pe acelasi plasmid, prin fenomenul transpozitiei repetate de material genetic. Plasmide de rezistenta au fost evidentiate la genurile Escherichia, Salmonella, Shigella, Proteus, Providencia, Klebsiella, Serratia din familia Enterobacteriaceae, la genurile Pseudomonas, Acinetobacter, Vibrio, Yersinia, Pasteurella, Campylobacter, Haemophilus, Neisseria, Bacteroides, Staphylococcus, Streptococcus, Bacillus, Clostridium, si Corynebacterium. Plasmidele prezente la bacilii gram-negativi sunt mai mari decat cele evidentiate la bacteriile gram-pozitive.
Plasmidul F se mai numeste si plasmid de sex, sau factor de fertilitate si contine, pe langa alti determinanti genetici, genele de transfer tra. Plasmidul F se poate transmite altor celule bacteriene prin conjugare. El se poate integra in cromozomul bacterian putand media transferul de gene cromozomiale de la o celula bacteriana donor la cea receptor. In functie de prezenta factorului F, bacteriile se impart in:
. bacterii F- lipsite de factorul F, denumite celule femele si care sunt receptoare de material genetic,
. bacterii F+, masculine, care au factorul F+, autonom, ca plasmid in citoplasma si care sunt celule donoare,
. bacterii Hfr care au factorul F+ integrat in cromozom, de asemenea masculine,
. bacterii F ' care au factorul F+ ca plasmid autonom, dupa ce acesta a fost integrat in cromozom si l-a parasit rupand un fragment ADN din cromozom. i aceste celule dunt donoare, deci masculine.
Bacteriofagii
Sunt virusuri care pazariteaza bacteriile. Se cunosc 6 grupe morfologice de bacteriofagi, cei mai bine studiati fiind bacteriofagii T ai bacilului coli.
Morfologie. Bacteriofagii sunt formati dintr-un cap hexagonal, un gat si o prelungire numita picior (coada). Capul este alcatuit dintr-un invelis proteic caracteristic virusurilor (capsida) si adaposteste ADN. Coada este un cilindru rigid invelit intr-un manson proteic asemanator miozinei si se termina cu o placa hexagonala ce contine o enzima de tipul lizozimului. De placa bazala se aprind 6 fibre cu rol in fixarea bacteriofagului pe suprafata bacteriei.
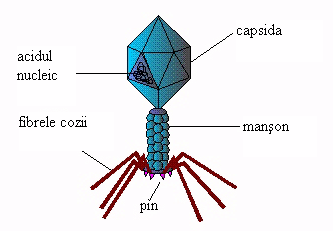 Atasarea bacteriofagului pe
suprafata peretelui bacterian este determinata de existenta unor
receptori de perete, specifici. Aceasta specificitate este de tip
enzimatic si sta la baza lizotipiei Dupa atasare are
loc contractia mansonului proteic si bacteriofagul isi
injecteaza numai ADN in celula bacteriana.
Atasarea bacteriofagului pe
suprafata peretelui bacterian este determinata de existenta unor
receptori de perete, specifici. Aceasta specificitate este de tip
enzimatic si sta la baza lizotipiei Dupa atasare are
loc contractia mansonului proteic si bacteriofagul isi
injecteaza numai ADN in celula bacteriana.
Dupa patrunderea genomului fagic in celula bacteriana, acesta va determina sinteza de noi bacteriofagi identici cu cel de la care a provenit ADN. ADN se replica prin replicare semiconservativa iar ribozomii bacterieni vor sintetiza proteinele capsidale si ale cozii. Dupa asamblarea noilor virusuri ele vor parasi celula bacteriana care se lizeaza. Acesta este ciclul litic al bacteriofagilor, iar ei se numesc fagi virulenti.
Dar nu intotdeauna relatiile bacteriofag-bacterie evolueaza in acest fel. Uneori genomul bacteriofagului se va integra in cromozomul bacterian incadrandu-se si functional in acesta. In aceasta situatie el nu se mai replica decat in acelasi timp cu cromozomul bacterian, deci in timpul diviziunii bacteriene si se numeste profag. Acest ciclu este ciclul lizogen iar bacteriofagii se numesc fagi temperati.
Lizogenia are urmatoarele consecinte pentru bacteria in cauza:
. bacteria lizogenizata este imuna la infectia cu acelasi bacteriofag, dar nu pentru alti bacteriofagi
. profagul codifica el insusi unele caractere pe care le dobandeste astfel bacteria lizogena. Un exemplu clasic in acest sens este toxigeneza la bacilul difteric care este codificata de un profag ce se afla in cromozomul bacterian. Tulpinile de bacili difterici care nu sunt lizogenizate de acest profag, nu sunt capabile sa secrete toxina si sunt, deci nepatogene,
. transductia, mecanism de transfer genetic de la o bacterie la alta, mediata de bacteriofagi si asupra careia vom reveni la variabilitatea bacteriana.

Figura 23 - Structura secventelor de insertie
3. Elemente genetice transpozabile
Elementele genetice transpozabile sunt fragmentele de insertie (IS) si transpozonii (Tn).

Figura 24 - Structura tranpozonilor
IS si Tn sunt fragmente mici de ADN cu limite structurale bine precizate care se pot integra repetat in mai multe situsuri dintr-un genom. Existenta lor a fost presupusa inca in anul 1931 si in anul 1952 de Barbara Mc Clintock care le-a denumit "gene saltarete" (jumping genes). Descoperirea a fost primita cu mult scepticism si neincredere la vremea respectiva, dar timpul a dovedit importanta ei, autoarea fiind onorata cu premiul Nobel in anul 1983.