|
|
|
|
Mecanismele activarii LT
Activarea LT, care reprezinta momentul central al majoritatii raspunsurilor imune fata de Ag timodependente NU se realizeaza in orice ipostaza, ci doar daca sunt strict indeplinite anumite conditii ce sunt diferite in vivo in vitro.
Conditii in vivo
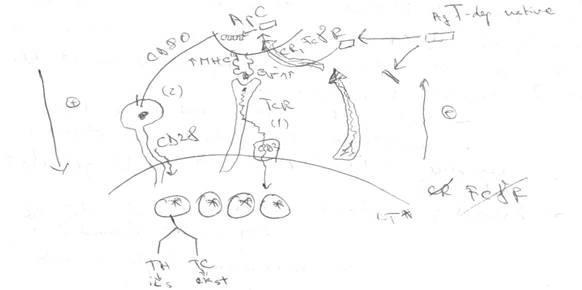
- Ag timodependente native NU pot stimula direct LT pentru ca LT NU dispun pe membrana lor de receptori de captare Ag.
- LT NU expun receptori pentru complement si IgG
- Exista alte celule (APC) care prezinta pe mebrana lor din abundenta receptori de captare Ag. Ele le capteaza, endociteaza, scindeaza => epitopi. Unii sunt ulterior prezentati in complex cu moleculele MHC. Ei sunt apoi recunoscuti de LT prin TCR+CD3
Distingem in cadrul cooperarii 2 tipuri de activare:
se desfasoara dinspre APC spre LT si la finele ei se realizeaza activarea LT. Se numeste activare anterograda pentru ca ea se exercita in acelasi sens cu prezentarea si respectiv recunoasterea antigenica
prin transmiterea unor semnale catre LT pe 2 cai principale:
primul semnal e transmis pe calea principala (TCR - CD3). El ia nastere ca urmare a recunoasterii de LT a epitopului prezentat. El are 2 roluri foarte importante:
1. conduce la activarea metabolica a LT
2. contribuie la declansarea diviziunii mitotice a LT => creste numarul de limfocite T per clona activata => 2 modificari clonale: activarea clonala si expansiunea clonala
cale accesorie: implica activarea receptorului CD28 prezent pe suprafata LT. Ele activat daca angajeaza legaturi cu un contraligand prezent pe APC proliferate (CD80). In CD28 => semnale transmise catre limfocit. Ele:
1. contribuie la activarea diviziunii mitotice a LT
2. contribuie la maturarea functionala a LT
3. contribuie la maturarea functionala a clonei
clona sufera 3 modificari foarte avantajoase:
1. activare
2. expansiune
3. maturare
Cu o asemenea clona => va declansa un RI eficient
dinspre LT deja activate catre APC
la finele ei => activarea totala, definitiva, maxima a APC
ea se exercita in sens opus prezentarii si recunoasterii Ag
se realizeaza prin eliberarea de catre LT activate a unor IL cu rol activator pentru APC (IFNγ, IL-4). Ele stimuleaza productia si expresia pe membrana APC a moleculelor MHC II (in special) => cresterea prezentabilitatii Ag => activarea suplimentara a LT
LT acivate suplimentar activeaza suplimentar APC, etc
Ele se activeaza reciproc pana in momentul in care este atinsa si depasita valoarea prag a declansarii RI
Valoarea prag a declansarii RI = atingerea si depasirea cand clona sufera cele 3modificari: e activata, expansionata si maturata
Activarea anterograda
trece intotdeauna prin 2 etape:
1. Etapa Ag nespecifica a activarii LT
in cursul ei se realizeaza legatura intre membranele APC si LT prin cuplurile de adeziune intercelulare
primele legaturi se realizeza pe seama cuplului de adeziune de linia I (CD2 pe LT si LFA-3 - CD58 - pe APC). Aceste legaturi sunt stabile pentru lungi perioade de timp. Numarul lor este imens de la bun inceput (sute de mii de asemenea legaturi)
apoi se stabilesc legaturi de linia a II-a, intre LFA-1 (pe LT) si ICAM. Ele apar mai tarziu datorita faptului ca cei 2 liganzi au o afinitate in general mica. Ele sunt instabile si initial numarul lor este mic.
Functiile acestei etape:
Prin formarea cuplurilor de adeziune membranara APC si LT vin in contact intim pe suprafete foarte mari
Conteaza pentru pregatirea activarii LT. In momentul in care se stabilesc legaturi CD2 - LFA-3, PLC, care in cazul LT e pozitionata mai interior, vine in contact cu CD2 => migreaza la suprafata, unde poate fi activata
ea are loc permanent. Legaturile NU au loc NUMAI in situatia in care APC prezinta un Ag, dar el NU a fost inca recunoscut. Ea are loc si in situatia in care APC NU expune epitopi (moleculele MHC ale APC sunt goale)
faptul ca LT stabilesc contacte continue cu APC => LT controleaza permanent APC daca expun sau NU Ag => supraveghere imunologica
2. Etapa Ag specifica a activarii LT
e ocazionata de prezentarea si recunoasterea Ag
epitopii sunt recunoscuti de TCR
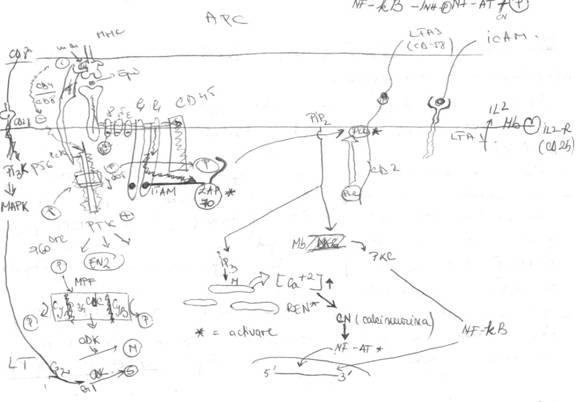
la TCR se asociaza laturile CD3 γ, δ si ε si la ele se asocialza cel mai frecvent 2 lanturi lungi, identice intre ele, lanturile ζζ. Apoi se mai asociaza, tot prin punti disulfurice, CD45
2 coreceptori:
CD4, daca e TH
CD8, daca e TC TS
CD4 CD8 se afla in citoplasma in contact cu o enzima, PTK (varietatea p56 LCK). In centrul catalitic exista o timozina, in pozitia 505, cu rol reglator pentru PTK
Activarea LT e rezultatul declansarii in succesiune a 2 categorii de stimuli:
1. Apare pe calea CD4 CD8. El e generat in momentul in care CD4 CD8 angajeaza legaturi cu domeniul monomorf al moleculei MHC => stimul ce se transmite prin CD4 CD8, el fiind un stimul inhibitor. El se transmite prin segmentul intracitoplasmatic al CD4 CD8 catre PTK. Primind stimulul, PTK sufera modificari conformationale => exteriorizarea Tyr din pozitia 505. Exteriorizata, ea se autofosforileaza spontan, ceea ce duce la inhibitia TOTALA a PTK. Inhibitia are rolul unui mecanism de protectie, prin care este impiedicata activarea LT in situatia in care APC NU prezinta nici un epitop (moleculele MHC sunt goale). Daca NU ar exista, LT ar reactiona fata de MHC si ar exista sanse ca LT sa distruga APC. Daca MHC NU sunt vide, atunci e declansat un al 2-lea semnal
2. apare pe calea TCR - CD3. El e un semnal activator si se transmite din lant in lant de la TCR prin toate lanturile CD3. atinge lanturile ζζ => urmeaza 2 cai: una scurata (a) si una lunga (b)
a. de la lanturile ζζ catre CD45. cd 45 se activeaza si incepe sa functioneze ca o PTP (fosfataza). CD45 preia (P) de pe Tyr din pozitia 505 => o defosforilare a PTK p56lck. Acum ea se activeaza. Ea fosforileaza enzime => activarea metabolismului intermediar al LT. In al doilea rand, ea fosforileaza secventele ITAM. Fosforilandu-le, ele se activeaza si devin capabile sa fosforileze alte enzime (kinaze). Cea mai importanta e kinaza ZAP-70. ea se fixeaza de lanturile ζζ (ZAP = Proteina asociata lanturilor ζζ, are GM de 70 kda). Stimulii sunt trimisi pe calea b.
b. De-a lungul lanturilor ζζ si in final au acces la ZAP-70, care tocmai s-a fixat. ZAP-70 se activeaza si fosforileaza preluand fosfatul de pe CD45 si il trimite la PLC, care fiind fosforilata, se activeaza. In mare, activarea declanseaza cascada fosfatidil-inozitolilor. PLC hidrolizeaza PIP2 (fosfatidil inozitol disfosfat) => Inozitol trifosfat (IP3) si diacilglicerol (DAG)
IP3
substanta hidrosolubila
difuzeaza in intreaga citoplasma, pana ajunge la nivelul REN, care prezinta receptori specifici pentru IP3 => activarea REN cu eliberarea masiva in citoplasma a Ca2+ (hipercalcitie). Efectul ei foarte important e acela ca Ca2+ activeaza calcinemina (o fosfataza calmodulin dependenta). Ea influenteaza indirect anumite functii nucleare. Ea exercita aceste efecte printr-un factor transcriptional = NF-AT (factor nuclear al LT activate). NF-AT e un factor polipeptidic prezent in mod continuu in citoplasma LT, dar atata timp cat LT se afla in repaus NF-AT e inactiv pentru ca e fosforilat. Activarea NF-AT se face prin defosforilalrea sa sub actiunea calcineminei. Defosforilat, NF-AT trece in nucleu si aici activeaza marea majoritate a genelor pentru IL => o sinteza crescuta de IL => naturarea LT
DAG
substanta liposolubila
NU difuzeaza in citoplasma, ci ramane la nivelul membranei
Aici DAG activeaza o kinaza (PKC). Ea, indirect, poate activa anumite functii nucleare prin intermediul unui factor transcriptional (NF-KB = factorul nuclear pentru lanturile usoare K ale Ig a LB)
NF-KB a fost evidentiat acum 20 de ani, in cazul LB mielomatoase
NF-KB - factor polipeptidic prezent in mod continuu in citoplasma LT. Cand sunt in stare de repaus NF-KB e inactiv, fiind cuplat cu un inhibitor proteic. Activarea se realizeaza sub actiunea PKC, care fosforileaza inhibitorul => desprinderea lui de pe NF-KB. NF-KB devine liber si poate trece in nucleu. Activeaza marea majoritate a genelor pt IL sau CK si in mod cu totul special activeaza foarte intens 2 gene: gena pentru IL-2 cat si gena pentru receptorul IL-2
LT devine producator de IL-2 si are si cu ce sa o recepteze => se activeaza cu prpria IL-2 => o activare autocrina cu IL-2 => activarea metabolismului si maturarea LT
Stimularea diviziunii mitotice
rezultatul transmiterii unor semnale pe 2 cai: TCR-CD3 si calea CD28
factorul cel mai implicat in stimularea mitozelor e MPF (mitosis promoting factor)
MPF-ul e un heterodimer format din 2 molecule de ciclina B, asociate la o componenta kinazica, reprezentata de P34cdc (cell division cycles). Activarea se realizeaza prin cuplarea unor fosforilari si defosforilari
Defosforilarea sub actiunea CD45, care defosforileaza ciclinele B => ele se desprind de pe kinaza
In paralel kinaza e activata prin defosforilare de o alta PTK, reprezentata de P60src (sarcoma cells). P60src se fixeaza de lanturile scurte ale CD3
Odata activata MPF sunt activate kinazele ciclin dependente CDK care actioneaza in punctele de restrictie ale ciclului mitotic situat intre faza G2 si faza M => aceste celule au toate sansele sa patrunda in mitoza
In faza G2 ajung ca urmare a transmiterii unor semnale pe calea CD28. ele apar cand CD28 angajeaza legaturi cu CD80 => semnale activatoare in CD28 ce se transmit la o kinaza asociata la CD28 si care se numeste fosfatidil inozitol 3 kinaza (PI3K). Ea activeaza cascada MAP kinazelor (proteinkinazele asociate mitozelor). MAPK stimuleaza kinazele ciclindependente responsabile de trecerea celulelor din faza G1 si S a diviziunii mitotice (punctul 2 de restrictie) => celulele ajung in faza M
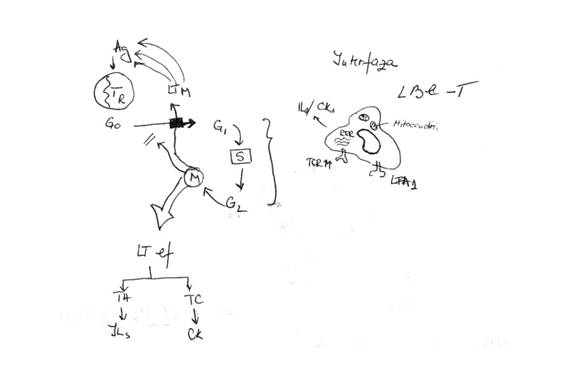
Consecintele morfofunctionale ale activarii LT
Activate, LT isi modifica atat aspectul mologic, cat si functia
LT in stare de repaus
Activat antigenic, trece din faza G0 in faza G1, apoi faza S in faza G2
LT din G1-S-G2 se afla in interfaza
Ele devin mai mari, cu aspect neregulat si prezinta o citoplasma foarte abundenta
In citoplasma creste numarul de mitocondrii => activarea metabolica a LT; creste RER => produc IL, CK; se modifica si expresia fenotipica: expresia de TCR pe suprafata lor (capacitate mai mare de a recunoaste Ag prezentate), creste expresia de LFA-1 (creste capacitatea de aderare a LT la APC)
In faza M marea majoritate a limfocitelor rezultate in urma diviziunilor devin LT efectorii (TH, producand IL sau TC, producand CK)
O mica parte raman in rezerva = LT cu memorie. Ele NU mai revin niciodata in faza G0 a ciclului mitotic, ci raman intr-o etapa intermediara, intre G0 si G1. Ele raman in stare preactivata. La un nou contact cu Ag, acestea vor raspunde foarte rapid.
Limfocitele B
Caractere generale
aproximativ 15-20% din totalul limfocitelor circulante
distribuite primar in ariile bursodependente ale organelor limfoide
celule putin recirculante
au durata de viata in general scurta (luni de zile)
Receptori de suprafata
3 categorii:
v Receptori de recunoastere Ag
v Receptori cu rol accesor in activare (CD45, CD25 sau IL-2R)
v Receptori de adeziune intercelulara (LFA-1)
I. Receptori de recunoastere Ag
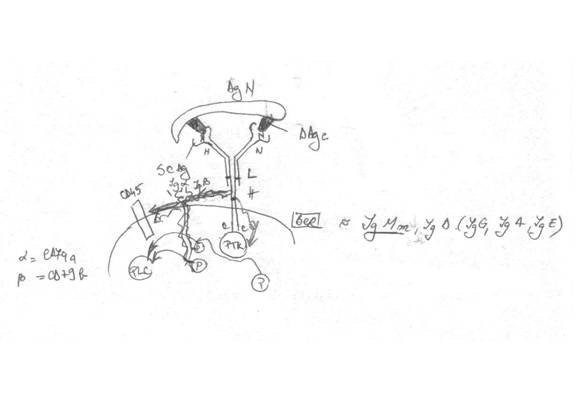
recunoasterea Ag se realizeaza printr-un complex receptorial, format din 2 componente:
v BCR (Ig transmembranare IgM forma monomerica, IGD, IgG, IgA, IgE). El e format din asocierea a 2 perechi de lanturi ( 2 grele - H - orientate cu capetele C-terminale in citoplasma). Prin capetele C-terminale se afla in legatura cu o PTK. Lanturile usoare (L) sunt pozitionate extramembranar
- aceste lanturi la capetele N-terminale se vor asocia intre ele si formeaza 2 cavitati = situsuri combinative pentru Ag (SCAg), prin care pot fi recunoscuti determiantii Ag conformationali prezenti la suprafata Ag nativ.
- desi are 2 SCAg, ele sunt identice intre ele => e recunoscut 1 singur determinant Ag => BCR e receptor monospecific
Functiile BCR:
1. recunoasterea Ag
recunoaste de regula Ag native aflate in stare solubila
in cazul LB NU e necesara niciodata prezentarea Ag. Ele singure pot sa-si capteze Ag prin BCR la care se asociaza receptorul de captare CR si FcγR
2. generarea unor stimuli activatori
acestia NU pot fi trimisi de BCR, de aceea se asociaza inca 2 lanturi cu functie de transductie a semnalului activator (Igα si Igβ)
De fapt ele sunt lanturi unice care in segmentul extracelular prezinta cate un domeniu pozitionat excentric
Actualmente Igα = CD79a si Igβ = CD79b
Ele se asociaza la BCR prin punti disulfurice, ca si la CD45
v CR si FcγR
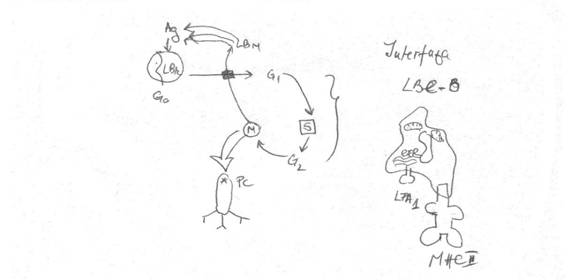
Activarea LB
mult mai complexa decat activarea LT
activarea se datoreaza trnsmiterii semnalelor activatoare pe 2 cai:
1. scurta
de la BCR la PTK
activandu-se, PTK preia (P) din citoplasma si il trimite catre diferite substrate (segmentele intracitoplasmatice ale Igα si Igβ, care sufera modificari conformationale ce permit trecerea unor semnale activatoare pe calea lunga
2. lunga
la CD45, care se activeaza si preia (P) de pe Igα si Igβ si il trimite la PLC => activarea PLC si declansarea cascadei fosfatidil-inoyitolilor
Consecintele morfofunctionale ale activarii LB
LB se transforma si morfologic, dar mai ales functional
LB trec din faza G0 in faza G1, apoi din S in G2
Ele trec in interfaza, unde devin niste limfoblasti B, care sunt mari si cu aspect neregulat si prezinta o citoplasma foarte abundenta ce inconjoara nucleul
In citoplasma gasim mitocondrii abundente si RER
Pe membrana creste expresia de LFA-1 si expresia de MHC II!!! Ele devin niste APC!!!
In faza M incep sa se divida. Cea mai mare parte devin plasmocite (produc Ac). O mica parte ramane in rezerva sub forma de LB cu memorie (raman intr-o etapa intre G0 si G1). La un nou contact ele vor actiona mai repede (memorie scurta pentru ca LB mor repede)
Organizarea sistemului imun
Distingem 2 componente:
1. componenta centrala
2. componenta periferica
Componenta centrala
reprezinta sectoarele de repartitie primara a tuturor clonelor limfocitare
reprezinta sectorul tranzitat in mod continuu de APC
reprezinta sectorul in care poate avea loc cooperarea APC - limfocite
reprezinta sectorul in care limfocitele pot fi si activate
a. Splina
2 zone: pulpa rosie (fara functie imunologica) si pulpa alba (functie imunologica)
Pulpa alba: dispusa sub forma a 3 mansoane, dispuse in jurul unei arteriole centrale
Primul manson (PALS intern) - zona timodependenta, populata de LT dispuse omogen
Al 2-lea manson (PALS extern) - zona bursodependenta, populata de LB prezente in aglomerari celulare = foliculi limfoizi
A 3-a = zona marginata, populata mai ales de Mcf
Functii
v Genereaza RI fata de Ag circulante, fata de care pot apare RIU, care se desfasoara integral in splina, sau RIC, care sunt doar initiate in splina si se desfasoara in tesutul Ag
Etapele desfasurarii RIU intrasplenice
- declansate de Ag circulante care pot fi prezente in circulatie ca Ag solubil sau Ag transportat
- Ag ajunge sub ambele forme in pulpa alba
1. Ag solubil
- mai repede ajunge sub forma de Ag solubil si patrunde in toate cele 3 zone, declansand RI doar in zona bursodependenta (Mcf NU au capacitatea de a capta Ag solubile, LT NU pot capta Ag)
- Ag e recunoscut de LB, ele se activeaza si se transforma in limfoblasti B. Dupa activare celula incepe sa se indeparteze de contactul antigenic, ramanand in PALS extern
2. Ag transportat de CDS
- CDS se afla in PALS intern, unde se transforma in IDC
- prezinta Ag LT prezente aici
- daca Ag e recunoscut, LT se activeaza si se transforma in limfoblasti T. Dupa un anumit interval de timp, celula se desprindede la locul cooperarii si migreaza, insa migrarea e foarte pronuntata si de regula limfoblastii T ajung in PALS extern, unde vin in contact cu limfoblastii B si incep sa coopereze
- consecinte: activarea completa a limfoblastilor B, care se transforma in plasmocite. Dupa un interval de timp ele migreaza si la distanta mare de cooperare => Ac ce sunt eliberati
b. Ganglionii limfatici
3 zone in parechim:
v Corticala (Cx) bursodependenta LB distribuite in aglomerari celulare = foliculi limfoizi. Care NU sunt stimulate => foliculi lifoizi primari
v Paracorticala (Pc) LT distribuite omogen
v Medulara celularitate mixta (LT, LB, Mcf, Plasmocite, etc)
Functii:
v Locul generarii unor RI fata de Ag tisulare => 2 raspunsuri:
RIU - integral in ggl
RIC - doar initiat in ggl, in rest se desfasoara in tesutul Ag
Etapele intraganglionare ale RIU:
declansat ca urmare a drenarii prin limfaticele aferente a unor Ag tisulare. Ele stimuleaza LB din foliculi, care isi modifica aspectul, devenind foliculi limfoizi secundari (au dimensiuni mari, enorme si avanseaza catre Pc). Modificarile se datoreaza faptului ca LB stimulate se activeaza si se divid, iar pe de alta parte migreaza.
Foliculul are 3 zone:
Subcapsulara (cap) formata din LB mici (neactivate)
Centrala, formata din putine LB (zona clara). Ele sunt LB mici. Sunt putine pt ca LB stimulate au migrat de aici si au trecut in paracorticala
Zona densa, formata din limfoblasti (Pc). Limfoblastii din Pc raman in contact cu LT de aici si coopereaza => se transforma in plasmocite, care migreaza, ajungand cel mai frecvent in medulara, unde elibereaza Ac.