|
|
|
|
ONTOGENIA CEULELOR HEMATOPOIETICE
Hematopoieza implica doua procese:
1. Dezvoltarea progresiva a caracterelor structurale si functionale specifice unei anumite serii celulare (diferentiere si maturatie).
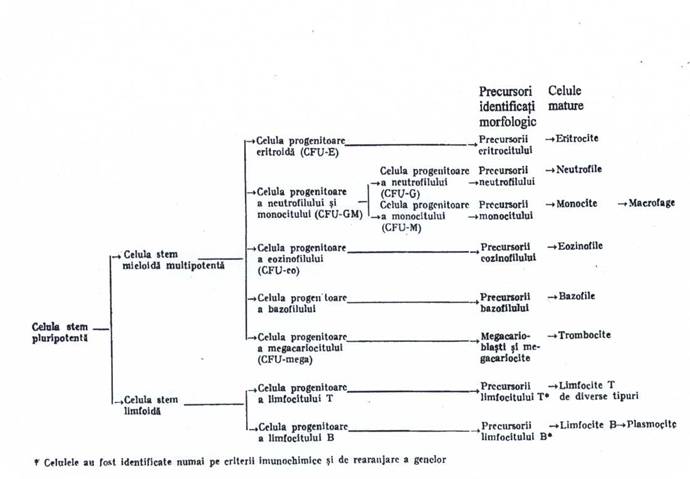
Celulele suse (stem) si celule progenitoare nu pot fi recunoscute morfologic pe frotiurile de maduva, dar pot fi studiate prin teste functionale. Celula hematopoietica primitiva, denumita si celula stem pluripotenta, va da nastere la doua tipuri de celule: celula stem meiloida multipotenta si celula stem limfoida. Celulele stem au capacitatea de a-si mentine constant numarul prin autoreinnoire ('ciclul de multiplicare'), alaturi de posibilitatea maturarii in alte tipuri celulare ('ciclul de maturare'). Celulele stem limfoide vor genera celulele progenitoare, care se vor matura in toate tipurile de limfocite: B, T si nule. Celulele stem mieloide multipotente se diferentiaza in variate tipuri de celule progenitoare mieloide, care vor urma linia de maturare in eritrocite neutrofile, eozinofile, bazofile si masrocite, monocite si trombocite. Spre deosebire de celulele stem (pluri. Si multipotente), celulele progenitoare limfoide si mieloide au o capacitate limitata de autoreinnoire.
Celulele pregenitoare unipotente, initiate in producerea unui singur tip de celula, sunt denumite celule formatoare de colonii (C.F.U.), deoarece in mediile de cultura, celulele progenitoare au capacitatea de a forma colonii micim in prezenta factorilor de crestere hematopoietici, specifici seriei (exemplu: eritropoietine). Aceste celule unipotente se vor matura in celule recunoscute morfologic pe frotiuri, care sunt denumite celule cap de serie: proeritroblasti, mieloblasti, monoblasti si megacarioblasti.
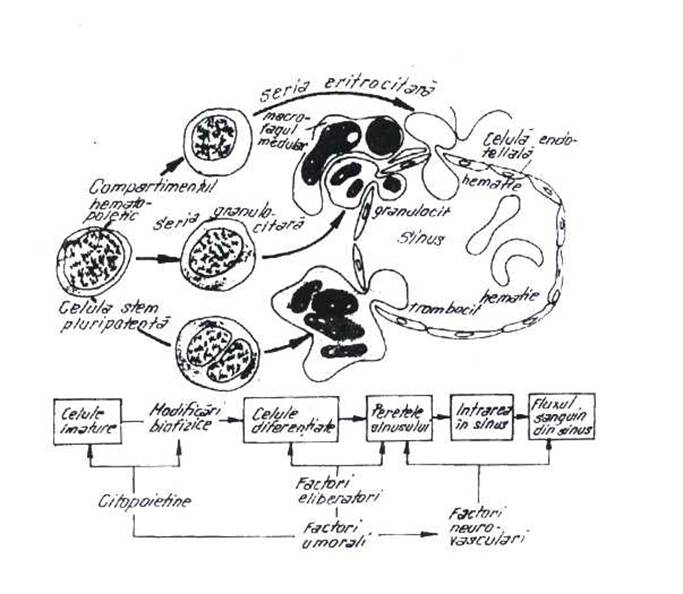
Fig. 1.1 - Structura maduvei osoase; insulele de hematopieza, maturarea si migrarea celulelor mature in sinusul medular.
Celulele hematopoietice derivate din celulelestem mieloide pot fi recunoscute morfologic.
Toate liniile mieloide urmeaza etapele de proliferare, diferentiere si maturare cu formarea in final a unor celule mature functionale, incapabile de diviziune.
Pentru fiecare serie mieloida, cu exceptia cele megacariocitare, au fost identificati, pe baza unor criterii morfologice si citochimice, precusori capabili de proliferare si maturare.
Pentru fiecare serie mieloida, cu exceptia celei megacariocitare, au fost identificati, pe baza unor criterii morfologice si citochimice, precursori capabili de proliferare si maturare. Seria megacariocitara este singura linie mieloida care isi pierde modelul de diviziune directa, dar isi pastreaza capacitatea de endomitoza (se divide numai AND-ul, iar citoplasma creste in dimensiuni).
In concluzie, hematopoieza poate fi descrisa dupa modelul unei piramide: varful este reprezentat de celula stem pluripotenta, iar baza, de celulele mature, identificate prin hemograma.
Eritropoieza
 Proeritroblastul
este o celula mare, cu o citoplasma redusa cantitativ, dar intens bazofila
(datorita prezentei unor numerosi ribozomi). Nucleul, de dimensiuni mari,
contine I cromatina fin dispersata si nucleoli vizibili.
Proeritroblastul
este o celula mare, cu o citoplasma redusa cantitativ, dar intens bazofila
(datorita prezentei unor numerosi ribozomi). Nucleul, de dimensiuni mari,
contine I cromatina fin dispersata si nucleoli vizibili.
Fig. 1.2 - Hematopoieza normala.
In etape succesive, proeritroblastul se diferentiaza in eritroblast bazofil, eritroblast policromatofil, eritroblast oxifil, ajungand in final in stadiul de reticulocit, care va parasi maduva osoasa si va trece in sangele periferic.
In cursul procesului de maturare, aceste celule nucleate vor suferi urmatoarele transformari:
a. reducerea progresiva a dimensiunilor nucleului si celulei;
In final, eritroblastul oxifil sufera un proces de expulzie a nucleului si se transforma in reticulocit medular. Reticulocitele formate in maduva intra in torentul circulator si raman in acest stadiu 1 - 2 zile, dupa care devin eritrocite mature.
Pe frotiurile de maduva si sange periferic, colorate May-Grünwald-Giemsa, reticulocitele sunt descrise sub forma unor celule rotunde, discret policromatofile (aspect bleu-cenusiu), cu un diametru mai mare decat hematiile adulte. Cand se foloseste coloratia supravitala cu albastru-crezil-briliant se observa precipitate bazofile, de aspect granular sau filamentar, formate din ARN ribozomal. Reticulocitele mai tinere prezinta precipitate de ARN sub forma unei retele bazofile, care a sugerat denumirea acestor celule. Hematiile adulte sunt lipsite de ribozomi.
Pe baza caracterelor morfologice, precursorii eritrocitari (eritroblastii) din maduva osoasa normala sunt denumiti normoblasti, iar procesul de formare a hematiilor adulte, eritropoieza normoblastica. Elementul major al eritropoiezei normoblastice este prezenta unei cantitati moderate de cromatina nucleara condensata in eritroblasti policromatofili.
Chiar si la persoanele sanatoase, un procent redus de eritroblasti (1-2%) parcurg anormal etapele de proliferare si maturare; ei vor fi recunoscuti si fagocitati de macrofagele medulare. Aceasta pierdere de eritrocite potentiale, ca urmare a distructiei intramedulare a precursorilor, este recunoscuta sub numele de eritropoieza ineficienta. Acest tip de eritropoieza devine important in transformarea megaloblastica a maduvei, ca urmare a eritropoiezei ineficiente se formeaza un pigment biliar timpuriu (early peak pigment), care poate fi identificat in stercobilinogenul fecal.
Granulocitopoieza neutrofilelor
Aparent, mieloblastii sunt morfologic asemanatori cu proeritoblastii, dar citoplasma lor este mai putin bazofila. Pana la aparitia granulocitului neutrofil circulant, mieloblastul parcurge succesiv stadiile de promielocit, mielocit neutrofiI, metamielocit neutrofil si granulocit neutrofil medular. Transformarile produse in cursuI acestui ciclu de maturare sunt urmatoarele:
1. reducerea progresiva a bazofiliei citoplasmei paralel cu marirea cantitatii de cromatina nucleara condensata, dupa stadiul de promielocit;
Megacariocitopoieza
In cursul formarii trombocitelor se realizeaza un model particular de mitoza (endomitoza), manifestat printr-o replicare a ADN-uiui, fara diviziunea nucleului si a celulei. Vor rezulta celule gigante uninucleate, cu 8-64 cariotipuri. Exista o corelatie stransa intre numarul de cariotipuri si gradul de lobulare a megacariocitului.
In cursul etapelor de proliferare si diferentiere ale seriei megacariocitare sunt recunoscute urmatoarele: tipuri celulare:
1. megacarioblastul - celula mare (15 μ), cu nucleu intens colorat, discret lobulat, cu citoplasma redusa si bazofilie marcata;
Megacariocitele trombocitogene sunt prezente in vecinatatea sinusoidelor medulare; la acest nivel, trombocitele se desprind progresiv din citoplasma megacariocitelor, printr-un proces de descuamare si trec in circulatie prin fenestratiile sinusurilor (fiecare megacariocit va genera 4000-6000 trombocite). La sfarsitul ciclului de viata, nucleul restant al megacariocitului va fi fagocitat de macrofage.
Monocitopoieza
Celulele sistemului monocit-macrofag sunt reprezentate pe linia maturarii de: monoblast, promocit, monocitul medular, monocitul din sangele periferic si macrofagul tisular.
Macrofagul tisular este celula functionala a acestei serii. Anumite macrofage, spre exemplu cele pulmonare, au capacitatea de a se divide local, asigurand astfel un numar adecvat de celule, pentru procesele de fagocitoza.
Limfocitopoieza
Celula stem limfoida din maduva osoasa genereaza celule progenitoare ale limfocitelor B, care vor suferi un proces de maturatie sub influenta micromediului din acest tesut. Vor rezulta limfocitele B, care vor trece in torentul circular si vor ajunge la nivelul organelor limfoide periferice, in zonele B.
Celulele progenitoare ale limfocitelor T, derivate tot din celula stem limfoida migreza in cursul procesului de limfopoieza din maduva osoasa in timus. La acest nivel, are loc maturarea precursorilor in limfocite T competente imunologic. . De asemenea, in timus se va face inventarul imunologic, in sensul ca limfocitele T care reactioneaza impotriva self-ului (antigenele proprii) vor fi inlaturate. In final, limfocitele T parasesc timusul, ajung in sange si, de aici, in organele limfoide, periferice, unde vor popula zonele T.
. Termenii folositii in denumirea celulelor corespunzatoare stadiilor de diferentiere a limfocitelor B si T sunt urmatorii:
o celula pre-B tanara. celula pre-B. celula B imatura. celula B matura.
Toate aceste celule prezinta caractere morfologice, fie de limfoblast, fie de limfocit. Identificarea diferitilor precursori limfocitari nu se bazeaza pe aspectul morfologic, ci pe urmatoarele criterii imunochimice:
o reactivitatea fata de anumiti anticorpi monoclonali;
Factorii stimulatori ai hematopoiezei
Factorii stimulatori ai hematopoiezei, denumiti si citokine, regleaza proliferarea, diferentierea si maturatia celulelor stem pluripotente, unipotente, durata de viata si functia celulelor mature hematopoietice.
In aceasta gama larga se inscriu:
o factorii stimulatori de colonii, cu denumirea internationala (C.F.S.);
Tabelul 1-3
Factorii stimulatori ai hematopoiezei
Factor
Sinonim international
Cromozom
Activitati biologice
Progenitori celule mature
S.F.
Factorul Steel
10
Activitate sinergica cu IL-3: GM-CSF, Epo asupra celulelor stem multipotente
IL-3
Multi-CSF
5q23-31
Celule stem multipotente:
eozinofil, bazofil
Continuare tabel 1-3
Factor
Sinonim international
Cromozom
Activitati biologice
Progenitori celule mature
G-CFS
17q11-21
Celule stem mieloide neutrofile
M-CFS
1p13-21
Celule stem mieloide monocite
Eritro-poetine
7q11-2
Celule pregenitoare eritroide
IL-5
E.D.T.
5q31
Celule progenitoare pentru eozinofile
IL-6
IFNα2
26kd-proteina
7p15
Lucreaza sinergic cu IL-3 si factorul Steel pe blasti
IL-1α
2q13
Introduce productia de factori stimulatori ai hematopoiezei
IL-1β
2q13
IL-4
BSF-1
5q23, 2-31;2
Lucreaza sinergistic cu eritropoietina G-CFS: M-CFS si progenitori
IL-8
Actiune pe neutrofil
(chematoxie)
IL-9
Colonii formatoare de serie eritroida
IL-10
Inhiba sinteza
Interferon - γ
IL-11
Sinergetic cu IL-3 cu factorul Steel pe celula multipotenta
IL-12
Suport pentru celule stem (cu IL-3 si factor Steel)
Induce sinteza
Interferon - γ
IL-13
2q23-31
Creste monocitele celule B-CD23, sinteza Ig si expresia clasa II antigen
Mpl
Trombopoetin
Are efecte pluripotentiale la nivelul celulei stem mieloide si a precursorilor megacariocitari
*Mpl - Ligand pentru factor de stimulare al trombocitului, capabil de a se lega de receptorul trombopoetinului. Numele provine de la 'murine mieloproliferative leukemia'.
Factorii stimulatori ai hematopoiezei au un domeniu extracelular, caracterizat prin una sau doua regiuni care contin 4 molecule de cisteina si complexul triptofanserina X triptofan-serina. Particularitatile care trebuie mentionate sunt urmatoarele:
o IL-6 contine in domeniul extracelular receptor pentru imunoglobina,
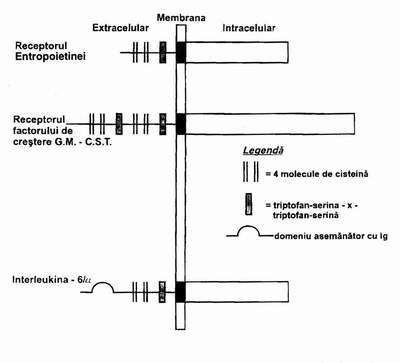
Fig. 1.3 - Diagrama unor factori stimulatori ai hematopoiezei.
Indicatii terapeutice
Cheia sustinerii hematopoiezei este celula pluripotenta. Prezenta acestor celule a fost demonstrata pentru prima data la soareci, prin experimentul lui Jacobsen si colab. (1949). Ei au aratat ca soarecii pot fi protejati de efectele unei iradieri totale (refacerea hemaatopoiezei), daca splina acestora este exclusa din campul de iradiere. Acest efect protector este sigur datorat unei celule, care provine din splina neiradiata. Caracteristica esentiala a celulelor stem imature este capacitatea lor de a repopula atat linia hematopoietica, cat si linia imuna. S-a dovedit astfel importanta functionala a celulelor stem.
Ulterior, Till si McCulloch (1961) au aratat ca in splinele soarecilor iradiati letal, dupa injectarea cu celule stem din maduva se formeaza celule hematopoietice functionale. Aceasta metoda a devenit o metoda cantitativa de apreciere a celulelor stem, international denumite CFU-S. Aceste colonii splenice (CFU-S) contin precursori ai eritroblastilor, ai seriei fagocitare se megacariocitare.
Numarul de celule stem este extrem de mic. Se apreciaza la 1 - 2,5 la 100.000 din celulele nucleate din maduva osoasa. Celule stem pot repopula atat sistemul mieloid cat si sistemul limfoid.
E.M. Kay (1965) a lansat ipoteza ca celule stem multipotente astazi marcate prin CD+34 contribuie prin formarea de clone, de progenituri, la mentinerea hematopoiezei in cursul vietii umane.
Facotrul de crestere al celulei stem multipotente este numir factorul Steel - CSF. El joaca rol in cresterea si dezvoltarea celulelor hematpoietice stem multipotente, a spermatocitelor, melanocitelor si a mastocitelor. Acest factor stimulator al celulei stem pluripotente contine 248 de amino-acizi, este codificat de o gena aflata pe cromozomul 10 la soarecii Steel. El reprezinta un ligand. Receptorul sau este numit C-Kit, este codificat pe cromozomul 5 din celula stem la soarecii numiti 'White spotting' (W). El este utilizat pentru mobilizarea celulelor hematopoietice progenitoare din sangele periferic (PBPC8). Factorul stimulator al celulei multipotente (CFS) lucreaza in armonie cu intereukina-3, interleukina-6, cu factorul de crestere al seriei granulocitare (G-CFS) si interleukina-11 la nivelul celului γ. Activitatea celulei stem CD 34+ (acestor factori), necesita ca celulele stem sa fie intr-o perioada activa a ciclului celular.
Factori stimulatori: G-CSF, GM-CSF, ai seriei granulocitare si monocitare au si ei o indicatie bine definita in tratamentul anemiilor aplastice, in aplazia medulara post-tratament sau in neutropeniile congenitale si/sau autoimune.
Reglarea hematopoiezei
Se presupune ca reglarea activitatii celulelor stem hematopoietice depinde de contactul intim cu una sau mai multe tipuri de celule stromale si de citokine din maduva osoasa. Proliferarea si maturarea celulelor pregenitoare intiate sunt influentate de o categorie speciala de factori de crestere, secretati de ceulel stromale (macrofage, celule encoteliale si fibroblasti); interleukina 3 (IL-3), factorul stimulator al coloniilor de macrofage si granulocite (GM-CSF), factorul stimulator al coloniilor de granulocite (G-CSF) si factorul stimulator al coloniilor de macrofage (M-CSF).
Rata eritopoiezei este controlata de eritropoetina, un hormon secretat de rinichi. Productia de eritropoetina este stimulata de scaderea aportului de oxigen la nivelul tesutului renal.
Acest hormon creste productia de hematii prin stimularea ratei de conversie a CFU-E in proeritroblasti. De asemenea, reduce perioada de timp necesara parcurgerii etapelor de diferentiere. De la proeritroblast la reticulocit medular si favorizeaza eliberarea reticulocitului in circulatie.
Reglarea proceselor de formare a celorlalte celule sanguine este inca insuficient cunoscuta.