|
|
|
|
STRUCTURA CELULEI BACTERIENE
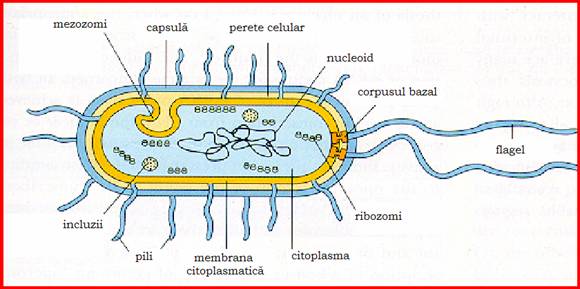
Figura 9- Structura celulei bacteriene
Unitatea morfofunctionala a bacteriilor este celula, un complex autoorganizat de molecule organice care schimba energie cu mediul inconjurator si este capabil sa-si regleze in mod autonom functiile vitale.
Bacteriile au o structura foarte complexa, fiind alcatuite din componente obligatorii, prezente la toate speciile bacteriene (nucleu, citoplasma, membrana citoplasmatica, mezozomi, perete celular) si componente facultative ce se gasesc doar la unele specii (capsula, flageli, fimbrii, spor).
1. Componentele obligatorii
1.1. Nucleul bacterian
Este un nucleoid sau echivalent nuclear, cu o structura primitiva in comparatie cu nucleul celulelor eucariote si reprezinta 2% din greutatea uscata a bacteriei. Nucleoidul este format dintr-o molecula circulara de ADN, organizata sub forma unui cromozom haploid care este in contact direct cu citoplasma datorita lipsei membrane nucleare. Molecula de ADN dublu spiralat este la randul ei suprahelicata in jurul unui miez de ARN, dispozitie necesara functionalitatii materialului nuclear.
In mod obisnuit, nucleoidul este situat in centrul celulei bacteriene si poate fi legat de membrana citoplasmatica prin intermediul mezozomilor.
Functia nucleului bacterian consta in depozitarea informatiei genetice necesara autoreplicarii, organizarii structurale si functionale a celulei bacteriene, deci a caracterelor ce definesc specia.
In afara de ADN cromozomial, la unele bacterii sunt prezente molecule circulare mici, extracromozomiale de ADN care se numesc plasmide si care se replica independent de cromozomul bacterian. Vom reveni asupra lor in capitolul de genetica bacteriana.
1. Citoplasma
Situata intre materialul nuclear si fata interna a membranei citoplasmatice, citoplasma este un sistem coloidal alcatuit din 80% apa, in care se gaseste o cantitate mare de molecule organice, ioni anorganici, enzime, ARN (de transport, mesager si ribozomial), vacuole si incluzii. Este lipsita de organitele celulare prezente la celulele eucariote cum sunt reticulul endoplasmatic, aparatul Golgi, mitocondriile, centrul celular, ergastoplasma.
La microscopul optic citoplasma apare omogena, amorfa, iar la cel electronic fin granulata datorita unui numar mare de ribozomi. Se apreciaza ca o celula bacteriana are 20.000 de ribozomi ce contin 80-90% din ARN-ul citoplasmatic.
Ribozomii sunt structuri sferice cu diametru de aproximativ 180nm si constanta de sedimentare de 70S. In absenta ionilor de Mg2+ se desfac in doua subunitati cu constanta de sedimentare de 50S si 30 S. Din punct de vedere chimic sunt alcatuiti din 65-70% ARN si 30-35% proteine. O parte din ribozomi se asociaza formand polizomi (mai ales in timpul sintezelor proteice), altii sunt liberi, iar o a treia categorie se ataseaza mezozomilor sau membranei citoplasmatice. Ribozomii reprezinta sediul sintezelor proteice din celula.
Incluziile citoplasmatice, descrise la unele specii bacteriene, sunt formatiuni structurale inerte, temporare, de diferite dimensiuni, variind in functie de specia bacteriana si conditiile de mediu. Compozitia lor chimica este diferita, ele putand fi de glicogen sau amidon (la unii bacilii aerobi sporulati), lipide (la genul Bacillus), polimetafosfati (sau incluziile de volutina descrise mai intii la Spirillum volutans si apoi de Babes si Ernst la bacilii difterici) etc. Incluziile sunt structuri legate de activitatea metabolica a celulei bacteriene si reprezinta un material de rezerva care poate fi folosit ca sursa de energie.
1.3. Membrana citoplasmatica
Este o membrana fina (6,5-7nm), elastica, lipsita de rezistenta mecanica ce margineste la exterior citoplasma bacteriilor si o separa de peretele celular. Pe sectiune apare trilaminata, fiind alcatuita din doua straturi fosfolipidice dispuse cu partile hidrofobe fata in fata. Printre moleculele fosfolipidice se gasesc molecule proteice, situate fie la nivelul unuia dintre cele doua straturi fosfolipidice, fie le traverseaza fiind expuse la ambele fete ale membranei. Aceasta structura dinamica, fluida, temporara si reversibila in functie de factorii de mediu este o "structura in mozaic", modelul ei fiind descris in 1972 de Singer si Nicholson.
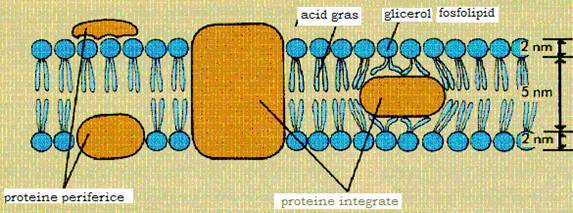
Figura 12 - Structura membranei citoplasmatice
Functiile membranei citoplasmatice:
este o membrana cu permeabilitate selectiva, indeplinind functia de bariera osmotica ce regleaza schimburile celulei bacteriene, in ambele sensuri cu mediul inconjurator. Permeabilitatea selectiva a membranei citoplasmatice permite realizarea in interiorul celulei, pentru unele substante, concentratii de 105 ori mai mari decat in afara celulei,
secreta numeroase enzime hidrolitice ce se elibereaza in mediul inconjurator unde scindeaza substratul nutritiv in unitati absorbabile,
unele proteine legate de membrana citoplasmatica joaca rolul de chemoreceptori,
indeplineste rolul mitocondriilor de la celulele eucariote, fiind sediul enzimelor lantului respirator si al fosforilarii oxidative, deci centrul energogenezei celulare,
este sediul sintezei acizilor grasi, a fosfolipidelor,
este implicata in sinteza peretelui celular, al polizaharidelor capsulare, participand activ la cresterea si diviziunea celulei bacteriene, la formarea sporului bacterian,
constituie o posibila tinta pentru actiunea unor chimioterapice ca, de pilda, polimixinele.
1.4. Mezozomii
Sunt structuri membranare care se formeaza prin invaginarea membranei citoplasmatice sub forma de buzunar sau in deget de manusa, prezente la bacteriile gram - pozitive si ocazional la cele gram - negative. Ei nu formeaza in citoplasma cavitati inerte, ci deschise spre spatiul periplasmic (spatiul dintre membrana citoplasmatica si peretele celular) si sunt in contact direct cu materialul nuclear.
Dupa morfologia lor, mezozomii sunt lamelari, veziculari si tubulari, iar dispozitia lor in celula bacteriana poate fi septala, periferica si nucleara.
Functiile mezozomilor:
participa la replicarea cromozomului bacterian si diviziunea celulara,
participa la reactiile de fosforilare oxidativa si oxidoreducere, dar in masura mai mica decat membrana celulara,
sediul unor enzime hidrolitice care indeplinesc rolul enzimelor lizozomale de la celulele eucariote,
sinteza si secretia unor exoenzime, ca de exemplu penicilinaza sau cefalosporinaza,
sinteza peretelui celular si formarea sporului bacterian.
1.5. Peretele celular
Componenta celulara obligatorie care inconjoara membrana citoplasmatica, peretele celular este o structura rigida, specifica bacteriilor. El este format dintr-un strat bazal, asemanator la toate bacteriile si un strat al structurilor superficiale, foarte diferentiat, in functie de care bacteriile manifesta caractere tinctoriale diferite: bacterii gram - pozitive, gram- negative si acido-alcoolorezistente.
Structura stratului bazal.
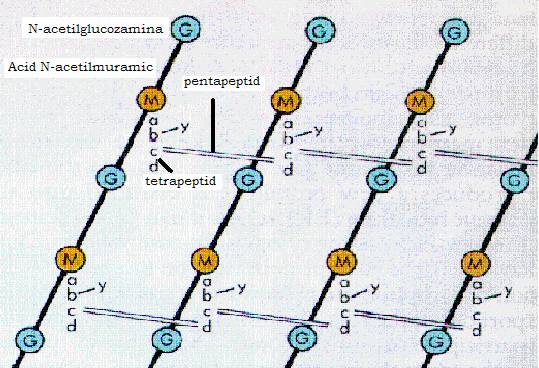
Figura 11 - Structura peptidoglicanului
Peptidoglicanul sau mureina este structura chimica responsabila de rigiditatea peretelui celular si care asigura forma si rezistenta mecanica a bacteriei. Prezent la toate bacteriile, el consta dintr-un schelet, format din molecule lungi paralele polizaharidice de N-acetil-glucozamina si acid N-acetil-muramic. Moleculele de acid N-acetil-muramic din lanturile vecine sunt legate intre ele prin punti polipeptidice, transversale, a caror structura difera la bacteriile gram- pozitive de cele gram-negative si chiar de la specie la specie. Astfel, de fiecare molecula de acid N-acetil-muramic se leaga un tetrapeptid ce contine D si L aminoacizi. De exemplu, tetrapeptidul la Staphylococcus aureus este format din L-alanina - D-glutamina - L-lizina - D-alanina, iar la bacteriile gram negative contine acid diaminopimelic.
La bacteriile gram - pozitive doua tetrapeptide de pe 2 lanturi paralele invecinate sunt legate printr-un pentapeptid. La bacteriile gram - negative, tetrapeptidele se leaga intre ele printr-o simpla legatura peptidica. Mentionam ca acidul diaminopimelic si acidul N-acetil-muramic sunt componente care nu au fost gasite in natura decat la bacterii.
Structura stratului structurilor speciale. Deosebim 3 tipuri de structuri speciale: gram-pozitiv, gram-negativ si acido-alcoolorezistent.
La bacteriile gram - pozitive, peptidoglicanul reprezinta 50-90% din greutatea uscata a peretelui celular, are o grosime de 15-30 nm si contine pana la 200 de lanturi paralele de mureina.
Stratul structurilor speciale este redus si alcatuit din polimeri hidrosolubili care sunt acizii theicoici: acidul ribitoltheicoic si gliceroltheicoic. Acestia pot patrunde pana la membrana citoplasmatica legandu-se covalent de aceasta - acizii theicoici de membrana sau numai pana la perete - acizi theicoici de perete. Ei reprezinta antigenele de supafata ale bacteriilor gram-pozitive.
Peretele celular al bacteriilor gram-pozitive este sensibil la actiunea lizozimului care rupe legaturile dintre acidul N-acetil muramic si N-acetilglucozamina si a penicilinei care inhiba sinteza peptidoglicanului.
La bacteriile gram-negative, peptidoglicanul are o grosime de 4-5nm, reprezentand numai 10% din greutatea uscata a bacteriei. Stratul superficial este insa mult mai complex decat la bacteriile gram - pozitive fiind alcatuit dintr-o membrana externa, lipoproteine si lipopolizaharidul de perete.
Membrana externa este formata dintr-un strat dublu fosfolipoproteic ce cuprinde o cantitate foarte mare de molecule proteice. Aceste proteine, bine definite la majoritatea speciilor bacteriene, sunt denumite Omp (outer membrane proteins) sau Momp ( major outer membrane proteins). Membrana externa se leaga de protoplast prin intermediul unei lipoproteine si a proteinei OmpA din membrana externa.
Proteinele nespecifice ale membranei externe cu functie de porine, cum sunt trimerii OmpF si OmpC, delimiteaza niste canale umplute cu apa prin intermediul carora pot difuza in spatiul periplasmic substante hidrosolubile.
Proteinele specifice au functii de transport pentru anumite substante, unele dintre ele fiind si receptori pentru bacteriofagi. De exemplu, proteina LamB este responsabila pentru transportul maltozei si receptor pentru fagul lambda al bacilului coli.
Proteinele de suprafata pot avea la unele bacterii rol in patogenitate, ca de exemplu cele 7 proteine de suprafata al shigellelor care le asigura invazivitatea.
Deasupra membranei externe a bacililor gram-negativi se afla lipopolizaharidul de perete (LPZ) sau endotoxina bacililor gram-negativi. Aceasta este alcatuita din:
lipidul A care are o structura particulara, fiind format din unitati dizaharidice de glucozamina, legate de beta-hidroxiacizi grasi cu 10-16 atomi de carbon fixati direct de membrana externa,
miezul sau "core", numit si antigen R, comun tuturor bacteriilor gram - negative si care include 2 zaharuri caracteristice: acidul keto-deoxyoctanoic si o heptoza.
unitati monozaharidice repetitive (15-40) care sunt specifice de specie si tip si constituie antigenul O al bacteriilor gram-negative.
LPZ este o toxina termolabila, care se elibereaza in mediul inconjurator de catre bacteriile gram-negative numai dupa liza lor si foarte reactiva in organismul gazda. Astfel, lipidul A produce febra, activeaza mecanismele apararii antiinfectioase si in exces produce socul endotoxic cu evolutie grava, chiar fatala.
Spatiul periplasmic. Sistemul structural dublu al peretelui celular la bacteriile gram-negative creeaza un compartiment ce se intinde de la membrana celulara pana la membrana externa, numit spatiu periplasmic. El contine peptidoglicanul si un gel care favorizeaza nutritia bacteriei prin continutul in enzime degradative ca, de pilda, fosfataze, nucleaze, proteaze etc. Tot aici sunt prezente enzimele de inactivare ale unor antibiotice cum sunt beta-lactamazele si cefalosporinazele.
Bacteriile acido-alcoolorezistente de interes medical sunt bacilul tuberculos si bacilul leprei a caror perete celular se aseamana cu cel al bacteriilor gram-pozitive. Structurile speciale contin acid micolic si o ceara ce confera acestor bacterii caractere tinctoriale deosebite si rezistenta crescuta la factorii de mediu. Astfel daca, dupa o incalzire de scurta durata ce topeste cerurile, un colorant patrunde in celula bacteriana, decolorarea acesteia sub actiunea acizilor sau alcoolilor nu se mai produce ca la alte bacterii. Acest caracter se numeste acido-alcoolo-rezistenta. Bacteriile acido-acoolo-rezistente se coloreaza foarte slab cu coloratia gram, evidentierea lor facandu-se la cald prin tehnica Ziehl-Neelsen.
Bacterii cu perete alterat sau formele L. Sunt bacterii cu stratul bazal viciat sub actiunea unor factori din mediu ca, de exemplu, lizozimul si penicilina, care lizeaza, respectiv impiedica sinteza peptidoglicanului.
Bacteriile lipsite total de perete celular se numesc protoplasti si provin din randul bacteriilor gram - pozitive, iar cele cu perete partial lezat, sferoplasti din bacteriile gram-negative, care isi pierd mai greu peretele.
Formele L sunt foarte sensibile la variatiile osmotice, putand supravietui numai in conditii speciale de osmolaritate, dar sunt foarte rezistente la antibioticele beta-lactamice. Daca factorii nocivi dispar din mediu, formele L se pot transforma in bacterii normale prin resinteza peretelui celular. Unele teorii mai vechi sustin ca formele L pot intretine si croniciza o infectie, dar dovezile in acest sens lipsesc.
Functiile peretelui celular:
asigura forma, rezistenta mecanica si osmotica a bacteriei;
asigura protectia membranei citoplasmatice fata de presiunea interna a celulei bacteriene, care este foarte mare: 5-6 atm la E. coli, 20-30 atm. la S. aureus;
regleaza traficul molecular de perete in ambele sensuri, deci schimbul de substante dintre bacterie si mediul inconjurator, fiind permeabil pentru molecule cu o GM mai mica de 10.000 daltoni si cu un diametru mai mic de 1nm;
stocheaza unele enzime in spatiul periplasmic la bacteriile gram-negative ce vor fi eliberate dupa necesitati, spre deosebire de bacteriile gram-pozitive care isi elimina enzimele direct in mediul extern;
prezinta receptori pentru bacteriofagi1;
este sediul antigenelor de suprafata, fiind deci implicat in raspunsul imun al macroorganismului;
este sediul unor factori de patogenitate;
are rol in diviziunea bacteriana si in procesul de sporulare.
Componente bacteriene neobligatorii
1. Capsula
Majoritatea bacteriilor patogene si comensale sunt capabile sa produca in vivo un material de invelis extracelular.
Capsula este un invelis compact, intim legat de celula bacteriana, cu o latime de cel putin 0,2µm. Este vizibila pe preparatele uzuale sau in coloratiile negative sub forma unui halou clar ce inconjoara bacteria.
Din punct de vedere chimic, capsula tuturor bacteriilor de interes medical este de natura polizaharidica (Streptococcus pneuomoniae, Klebsiella etc.) cu exceptia capsulei bacilului carbunos care este polipeptidica.
Capsula are rol in rezistenta bacteriilor fata de fagocitoza, fiind astfel un factor de virulenta. Variantele necapsulate ale acelorasi specii sunt nepatogene. De exemplu, Streptococcus pneumonie, de tip S, capsulat, produce la soarecele alb de laborator o septicemie mortala, pe cand varinata necapsulata nu este patogena.
Capsula este o structura cu proprietati antigenice specifice (antigenele K) care permit diferentierea unor serotipuri in cadrul speciei.
Bacteriile nu sintetizeaza material capsular in vitro pe mediile uzuale, coloniile rezultate fiind rugoase, de tip R ="rough". Pe medii speciale, insa, bacteriile pot creste capsulate, coloniile fiind mucoase, de tip S ="smooth".
Structura descrisa mai sus este "capsula clasica" a bacteriilor, la care ne referim atunci cand vorbim de bacterii capsulate. La unele bacterii s-au mai evidentiat si alte structuri de invelis, cum sunt:
Microcapsula este o structura discreta cu o grosime sub 0,2µm care nu se evidentiaza la microscopul optic, ci numai prin metode imunologice sau electronomicroscopice (Neisseria gonorrhoeae). Ea constituie un factor de virulenta.
Stratul mucos, glicocalixul este un strat amorf si vascos ce inveleste bacteria. El este format din lanturi lungi de polizaharide, cum sunt levanii si dextranii, cu rol major in adezivitatea bacteriilor de suprafete. Astfel, de pilda, Streptococcus mutans produce cantitati mari de dextran si levan prin intermediul carora se ataseaza de suprafata dintiilor contribuind la formarea cariilor si a placii dentare. Un alt exemplu este Pseudomonas aeruginosa, care secreta un strat mucos dens care ii creste rezistenta la antibiotice.
Functiile capsulei:
este un factor de aderenta si colonizare a bacteriile pe suprafete;
protejeaza bacteriile de diferiti agenti antibacterieni din mediu cum sunt: bacteriofagii, colicinele, complementul, lizozimul sau alte enzime bacteriolitice;
protejeaza bacteriile de actiunea fagocitelor, fiind deci un factor de virulenta;
reprezinta sediul antigenelor capsulare, importante in identificarea acestor bacterii.
Cilii sau flagelii bacterieni
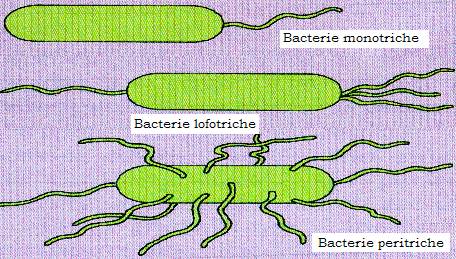
Figura 13 - Cilii bacterieni
Sunt apendici filamentosi ai speciilor bacteriene mobile, cu originea in citoplasma bacteriana, si servesc ca organe de locomotie. Ei sunt prezenti mai ales la bacili (Enterobacteriaceae) dar si la unii coci (enterococ). Cilii sunt structuri helicoidale, cu dimensiunea in general mai mare decat cea a celulei bacteriene careia ii apartin (2-15/2-20 nm) si se evidentiaza microscopic prin coloratii speciale.
Dispozitia si numarul cililor sunt caracteristice speciei. S-au descris bacterii atriche (fara cili), monotriche - cu un cil polar (Vibro cholerae), lofotriche - cu un smoc de cili situat la unul din polii bacteriei - (Pseudomonas fluorescens), amfitriche - cu cilii situati la ambii poli ai bacteriei (la genul Spirillum) si peritriche - cu cilii dispusi pe intreaga suprafata a bacteriei (Salmonella, E. Coli, Proteus etc.).
Structura cililor este tubulara, ei fiind formati dintr-o proteina contractila, flagelina, sub forma polimerizata. Cilul se ataseaza de bacterie printr-un corpuscul bazal si un carlig de articulatie aflate in citoplasma bacteriei. Corpusculul bazal se insera prin carligul de articulatie de membrana citoplasmatica si peretele celular.
Mobilitatea si directia miscarii cililor pot fi influentate de concentratia unor substante din mediu (chimiotactism pozitiv sau negativ) si de temperatura. Ei sunt sediul antigenelor flagelare (H), importante in identificarea bacteriilor.
Cilii bacterieni nu se observa decat pe preparate colorate special. In practica de rutina nu se evidentiaza flagelii, ci mobilitatea bacteriilor, fie prin examinarea lor pe un preparat nativ intre lama si lamela in care se urmaresc miscarile bacteriilor, fie prin insamantarea tulpinii pe un mediu semisolid prin inteparea in profunzime a mediului. Daca bacteria creste nu numai pe traseul insamantarii si difuzeaza in mediu, ea este mobila.
3. Pilii bacterieni
Numeroase specii bacteriene gram-negative au pe suprafata lor lor niste apendici filamentosi, rigizi, mai scurti decat flagelii, in numar mare (100-500/celula) si cu dispozitie in general peritricha. Ei se evidentiaza numai prin microscopie electronica.
Din punct de vedere chimic sunt polimeri proteici de pilina, dispusi helicoidal intr-o structura tubulara. Natura proteica a pililor le confera proprietati antigenice. Din punct de vedere functional, pilii se impart in:
sex pili - codificati de formatiunile genetice extracromozomiale (plasmide) si care prezinta o importanta deosebita in transferul de material genetic intre bacterii. Sunt prezenti mai ales la bacteriile gram-negative (Enterobacteriaceae, Pseudomonas),
pili comuni sau pili de aderenta (fimbrii), in numar mare, codificati cromozomial, si care servesc bacteriilor la fixarea ferma de mediul de cultura sau de celulele pe care se afla. Ei constituie, deci, un factor important de virulenta (de exemplu la gonococ). In afara de aderenta, pilii comuni mai au si proprietati antifagocitare.
Ambele categorii de pili prezinta antigene specifice piliare si pot fi receptori pentru bacteriofagi.
4. Sporii
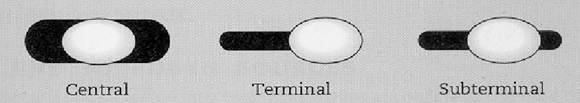
Figura 14 - Pozitia sporului
Unele bacterii se transforma in spori, care sunt forme primitive de diferentiere celulara, cu rezistenta crescuta la factorii de mediu si care apar endocelular in conditii nefavorabile de viata. Sporogeneza se intalneste numai la 3 genuri de bacterii gram-pozitive, Clostridium si Bacillus care sunt bacili si Sporosarcina care sunt coci.
Sporii nu sunt forme de inmultire ale bacteriilor. Dintr-o bacterie vegetativa se formeaza un singur spor, care in conditii favorabile de viata va da nastere unei singure celule bacteriene.
Forma sporului poate fi rotunda sau ovala. Diametrul sporilor este mai mic la bacilii sporulati aerobi, nedepasind diametrul bacteriei (genul Bacillus), pe cand la genul anaerob Clostridium, sporul are un diamentrul mai mare decat bacteria, producand deformarea acesteia. Acest caracter al sporului se numeste "caracter clostridial" si este important in identificarea bacililor gram-pozitiv anaerobi.
Pozitia sporului bacterian constituie un caracter taxonomic. Poate fi centrala, ca la bacilul carbunos (Bacillus anthracis), subterminala, ca la B.cereus, sau terminala, ca bacilul tetanic (Cl.tetani).
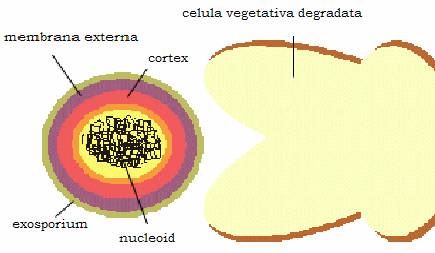
Figura 15 - Structura sporului
In coloratiile obisnuite sporul apare ca o zona incolora in corpul bacterian. El se evidentiaza insa prin coloratii speciale, la cald, care permeabilizeaza invelisurile sporale pentru coloranti (coloratia Möller). Pe preparatele native intre lama si lamela efectuate din culturi, sporii apar ca niste formatiuni rotunde, refringente.
Ultrastructura si compozitia chimica a sporilor asigura acestora o mare rezistenta la agentii fizici si chimici, ceea ce face ca ei sa fie considerati forme de rezistenta ale bacteriilor. De importanta deosebita pentru medicina este rezistenta sporilor la caldura, de care trebuie sa se tina cont la sterilizare.
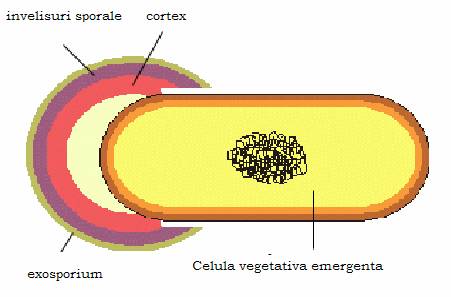
Figura 16 - Germinarea sporului
Sporularea. Este un proces ce se declanseaza in conditii nefavorabie de viata, cand bacteriile sunt lipsite de surse nutritive. Ea presupune formarea unor structuri noi si disparitia altor structuri caracteristice formei vegetative ale bacteriei.
Din punct de vedere morfologic, sporularea incepe prin condensarea la un capat al celulei a materialului nuclear. Acesta va fi separat de un sept ce provine din invaginarea membranei citoplasmatice. Septul creste, se desprinde de membrana citoplasmatica si formeaza in jurul materialului nuclear o membrana dubla a carei fete care se privesc sunt cele ce sintetizeaza peretele bacterian. Acesta este stadiul de prespor.Cele doua lamele ale membranei presporale vor sintetiza invelisurile sporale: peretele sporal si cortexul situate intre cele doua lamele ale membranei presporale si o tunica proteica si exosporiumul in afara membranei presporale.
Structura sporului variaza de la specie la specie, dar organizarea generala se aseamana. De la interior spre exterior, sporul este format din:
. core sau protoplastul sporal, format din materialul nuclear inconjurat de membrana citoplasmatica. Aici este depozitat DNA si o substanta care are rol in rezistenta sporului la caldura - dipicolinatul de Ca, care nu s-a mai evidentiat nicaieri in natura,
. cortexul intern sau peretele sporal format din peptidoglican si care este peretele celular primordial,
. membrana externa sau cortexul sporal, care este stratul cel mai gros al sporului si care contine si el un peptidoglican, cu o structura particulara, foarte sensibil la lizozim. Autoliza acestui strat este momentul cheie in transformarea sporului in forma vegetativa,
. tunica proteica este formata din proteine chitinoase cu numeroase legaturi disulfitice. Ea este impermeabila, fiind responsabila de rezistenta sporilor la unele dezinfectante,
. exosporiumul este prezent numai la unii spori si contine lipoproteine si zaharide.
Germinarea sporului. In conditii de mediu favorabile, sporul germineaza si va da nastere unei bacterii vegetative identice cu aceea in care s-a format. Germinarea are loc in 3 etape:
Activarea sporului. Se produce cand acesta intalneste conditii favorabile de viata, dar si un factor care sa lezeze invelisul sporal (mecanic sau chimic ca, de exemplu, lizozimul, compusi cu grupari sulfhidrilice libere etc.).
Initierea este declansata de un mediu nutritiv bogat. Pentru unii spori triggerul este L-alanina, iar pentru altele adenozina. Acesta duce la activarea unei enzime autolitice care degradeaza cortexul sporal. Are loc absorbtia de apa, eliberarea dipicolinatului de calciu si degradarea unor compusi sporali.
Dezvoltarea sporului. Protoplastul sporal se transforma in bacterie vegetativa, care trece printr-o perioada metabolic activa de refacere a constituentilor celulari normali si a echipamentului enzimatic complet.
3. STUDIUL MORFOLOGIC AL BACTERIILOR
Examinarea microscopica este frecvent primul pas in identificarea unor bacterii. Caracterele morfologice si tinctoriale incadreaza bacteriile intr-unul din grupurile majore stabilind conduita diagnosticului bacteriologic. Elementele morfologice importante in identificarea unei bacterii sunt: dimensiunea, forma, asezarea, prezenta unor structuri speciale ca, de pilda, endospori, flageli, capsula sau incluzii intracelulare.
Microscopul cu camp luminos. Acesta are 2 sisteme de lentile care maresc imaginea obiectului studiat: lentilele obiectivului si lentilele ocularelor. In mod obisnuit se folosesc 3 obiective: un obiectiv care mareste de 10x si care se foloseste pentru obtinerea unei imagini de ansamblu a preparatului examinat, un obiectiv ce mareste de 40x, cu care se examineaza microorganisme mai mari ca, de pilda fungi, paraziti si in fine obiectivul cu imersie care amplifica imaginea de 90 sau 100x si cu care se examineaza bacteriile. Imaginea mai este amplificata inca de 7 sau 10 ori de lentilele ocularelor. Puterea de rezolutie (distanta cea mai mica intre 2 puncte ce poate fi perceputa) a unui microscop fotonic este de 0,2µ. La microscopul cu camp luminos se examineaza preparatele colorate, dar uneori si preparate native.
Microscopul cu fond intunecat. Are un condensator special, cardioid, care permite patrunderea luminii numai de la periferia obiectivului. La acest tip de microscop se examineaza preparatele native, intre lama si lamela, efectuate direct dintr-un produs, in care se cauta bacterii a caror latime nu depaseste 0,1-0,2µ (ca, de exemplu, evidentierea treponemelor din sancrul sifilitic), sau dintr-un mediu de cultura lichid, in care se urmareste dezvoltarea unei culturi (Leptospira etc.).
Microscopul cu contrast de faza se foloseste tot la examinarea preparatelor native. El are un sistem de filtre care permit sesizarea diferentelor de faza a luminii cand aceasta trece prin obiecte de densitate diferita. Microorganismele examinate la acest microscop au aspect tridimensional.
Microscopul cu raze UV. Unii compusi ca, de exemplu, fluorocromii sunt capabili sa absoarba lumina ultravioleta si sa eminta o energie cu lungime de unda superioara. Preparatele de examinat sunt colorate in prealabil cu un compus fluorescent ca, cum este, izotiocianatul de fluoresceina iar apoi sunt examinate. Particulele colorate vor apare luminoase, fluorescente pe campul intunecat al microscopului.
Microscopul electronic. Lumina este inlocuita cu un fascicul de electroni. In microscopia electronica cu transmisie, electronii trec direct prin preparat, iar in microscopia prin baleiaj ei trec dintr-un anumit unghi prin preparat, astfel incat imaginea ce apare pe ecran este tridimensionala.
Examinarea morfologiei bacteriene se efectueaza pe diverse preparate microscopice.
Efectuarea frotiurilor fixate si colorate este o metoda de rutina in laboratorul de bacteriologie. Frotiurile se pot colora simplu cu un singur colorant, elementele preparatului avand toate aceeasi culoare, eventual de intensitate diferita, sau diferentiat cu mai multi coloranti, bacteriile colorandu-se diferit in functie de afinitatea lor tinctoriala.
Dupa efectuarea frotiurilor, acestea se usuca la temperatura camerei si se fixeaza cel mai des prin caldura, trecand frotiul de 2-3 ori prin flacara. In timpul fixarii, formele vegetative ale bacteriilor sunt omorate si devin permeabile pentru coloranti. Fixarea chimica (cu alcool metilic, acetona, May-Grünwald, acid osmic etc.) se utilizeaza mai ales cand preparatele sunt sectiuni de tesuturi sau frotiuri de sange, deoarece protejeaza celulele din produs de degradare.
Coloratiile simple folosesc solutii apoase a unui colorant bazic ca, de exemplu, albastru de metilen, violet de metil, fuxina bazica etc. Cationii colorantilor bazici se combina cu anionii protoplastului bacterian si cu fosfatii acizilor nucleici. Dupa colorare, frotiurile se spala pentru indepartarea excesului de colorant. Colorantii acizi nu coloreaza bacterii decat la un pH foarte acid, dar pot fi folositi pentru coloratii negative. Precizam ca peretele celular nu se coloreaza in mod obisnuit, structura colorata observabila la microscop fiind protoplastul bacterian. Spatiul dintre bacteriile dispuse in lanturi corespunde tocmai peretilor celulari invecinati necolorati. Unii microbi se coloreaza bipolar datorita dispozitiei citoplasmei la ambele capete ale bacteriei.
Coloratia negativa este o metoda rapida si simpla de studiu a morfologiei bacteriene. Bacteriile se amesteca cu colorantul (tus de China) si se intind ca un film pe lama. Bacteriile apar incolore pe fondul colorat al preparatului.
Impregnarea argentica se utilizeaza pentru colorarea spirochetelor, mai ales cand se urmareste evidentierea lor in tesuturi. In aceasta coloratie, spirochetele se coloreaza in brun-negru pe fondul galben al preparatului.
Coloratiile diferentiale sunt cele mai importante in laboratorul de bacteriologie. Cel mai des folosita este coloratia gram, introdusa de studentul danez Christian Gram la sfarsitul secolului trecut. Dupa cum apar bacteriile in aceasta coloratie ele se impart in gram-pozitive si gram-negative.
Coloratiei gram consta in colorarea initiala a bacteriilor cu cristal-violet sau violet de gentiana. Mordansarea se face cu lugol. Urmeaza decolorarea in urma careia unii microbi, cei gram-pozitivi, pastreaza colorantul violet, iar altii, cei gram-negativi, vor fi decolorati cu un amestec de alcool-acetona. Acestia din urma se vor recolora cu fuxina bazica. Bacteriile gram-pozitive vor aparea deci de culoare violet iar cele gram-negative rosii. Este important de mentionat faptul ca bacteriile isi pot schimba caracterele tinctoriale in conditii neprielnice de viata, cele gram-pozitive mai ales putandu-se decolora.
Proprietatea bacteriilor de a se colora diferit prin aceasta coloratie exprima diferente de structura a peretelui celular. Mecanismul care sta la baza acestei coloratii nu este inca elucidat.