|
|
|
|
GENETICA POSTMENDELIANA
1. Spectrul dominantei
1.1. Dominanta incompleta
o F1 nu mai apar caractere dominant si recesiv, ci unul intiermediar
o P - fenotip flori rosii + fenotip flori albe
o F1 - descendenti cu flori roz
o F2 - incrucisarea hibrizilor = segregare
flori rosii - flori roz - flori albe in raport 1 : 2 : 1
o F3 - prin autopolenizarea indivizilor din F2 au aparut:
fenotip flori rosii → descendenti cu flori rosii = genotip RR
fenotip flori albe → descendenti cu flori albe = genotip rr
fenotip flori roz → descendenti cu fenotip diferit
v rosu, roz si alb in proportie 1 : 2 : 1
v indivizii din F1 au fost heterozigoti Rr
v in F2 - segregarea alelelor in gameti conform legii I-a a lui Mendel
v reasocierea la fecundare = genotipuri RR (1/4), Rr (1/2) si rr (1/4)
v caracter intermediar intre dominanta completa - recesivitatea completa
v dominanta incompleta (dominanta partiala, semidominanta)
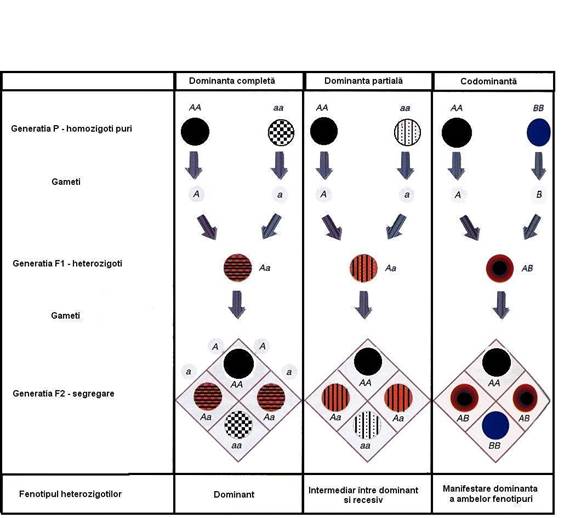
Figura 2-7. Spectrul dominantei.
1.2. Codominanta
A (1/4) ← genotip homozigot dominant IAIA
AB (1/2) ← genotip heterozigot codominant IAIB
B (1/4) ← genotip homozigot dominant IBIB
v raport de segregaare 1 : 2 : 1
exemplul - heterozigoti pentru anemie falciforma - genotip HbAHbS
o efectul de dominanta este diferit in functie de nivel:
de organism = dominanta completa a alelei HbA fata de alela HbS
v fenotip normal - subiect sanatos
celular = dominanta incompleta a alelei HbA fata de alela HbS
v asocierea fenotip celular normal - patologic
v discocit - siclemie, fara efect patologic
molecular = codominanta celor doua alele HbA si HbS
v se produc in cantitati egale ambele tipuri de hemoglobina
2. Alelismul multiplu
la un singur individ pot exista pentru un caracter cel mult doua alele
la indivizi diferiti pot exista pentru un caracter mai multe alele
o mai mult de doua variante alelice pentru o gena = serie alelica
o exemplul cel mai cunoscut
grupele sanguine: IA, IB si i
in cadrul alelelor IA exista o subserie alelica: IA-1 - IA-6
in cadrul alelelor IB exista o subserie alelica: IB-1 - IB-3
o alelele dominante - caracter normal = alele salbatece
larg raspandite ← avantajul selectiv
o alelele modificate (dominante / recesive: rare) = alele mutante
3. Interactiunea dintre gene
dominanta si recesivitatea = proprietati intrinseci ale alelelor
alela dominanta nu o inhiba pe cea recesiva
efectele fenotipice → confera caracterul de dominanta sau recesivitate
interactiunile genice = activitatea unei gene - influentate de o gena nealelica
o epistaziea = influenta unei gene nealelice asupra exprimarii altei gene
o pleiotropiea = influenta unei gene mutante asupra unor caractere diferite
exemplu - sindromul Marfan ← mutatie
v statura inalta-gracila / defecte cardio-vasculare / miopie
exemplu - fibroza chistica de pancreas
v mutatie CFTR ΔF 508 proteina transportoare de Cl-
v functia anormala a glandelor exocrine: plaman, ficat, pancreas, mucoase, piele sau gonade
penetranta variabila = lipsa fenotipului specific codificat de genotipul specific
datorita influentelor mediului sau dezvoltarii
o se refera la proportia de indivizi care exprima gena respectiva
o exemplu, nu toti purtatorii mutatiei pentru retinoblastom fac boala
expresivitatea variabila = exprimarea completa sau partiala a unei alele
o exemplu - sindromul Marfan
numai fenotipul - talie inalta si desirata
in plus o boala cardiovasculara saqu la unii este prezenta si miopia
o exemplu - fibroza chistica de pancreas
boala cu tabloul clinic complet, letala in prima copilarie
manifestare frusta, de exemplu o "simpla" sinuzita cronica
4.a. Teoria cromosomiala a ereditatii
Descoperirea cromosomilor
o ignorata pana in 1900
o nu a fost asociata cu legile lui Mendel
asocierea cromosomilor = factorii mendelieni - 1902, W. Suton si T.Boveri
o cromosomii perechi (omologi) - perechi de alele (genotip)
o omologii si alelele segrega in mod egal in gameti
o neomologii si perechile nealelice segrega in mod independent una de alta
diferiti cercetatori au emis ipoteza ca genele sunt localizate pe cromosomi
locus = pozitia unei gene pe un anumit cromosom
o loci = pozitiile diferitelor gene (pluralul de la locus)
Drosophyla melanogaster - prima specie cariotipata
o N.Stevens (1905) - existenta a 8 cromosomi
o 4 perechi omoloage la indivizii de sex feminin
o 3 perechi omoloage si una neomoloaga la indivizii de sex masculin
o perechile comune - cromosomi autosomi sau autosomi
o perechea diferita - cromosomi heterosomi (heterosomi sau gonosomi) sau cromosomi de sex
sexul feminin - perechea 4 (omologi) - cromosomi X
sexul masculin - perechea 4 = un cromosom X + cromosom Y
o determinismul cromosomial al sexelor → teoria cromosomiala a ereditatii
4.b. Teoria cromosomiala a ereditatii
- T.Morgan, in experimente pe Drosophyla melanogaster
corelare segregare cr. X - segregare genea → caracterul mutant: ochi albi
P - ♂ ochi albi (genotip XwY) + ♀ ochi rosii (genotip Xw+Xw+)
F1 - toti indivizii: ochii rosii (heterozigoti - genotip Xw+Y sau Xw+Xw)
o alela mutanta → culoarea alba a ochilor = recesiva
F2 fenotipul ochi rosii - ochi albi segrega in raport de 3 : 1
o toti indivizii cu ochi albi erau de sex masculin
o gena → culoarea ochilor - localizata pe cr. X - combinatii posibile:
genotip femel Xw+Xw+ → fenotipul ochi rosii
genotip femel Xw+Xw → fenotipul ochi rosii
genotip masculin Xw+Y → fenotipul ochi rosii
genotip masculin XwY → fenotipul ochi albi
F3 - ♂ cu ochi albi + ♀ (descendentele lor - heterozigoti) cu ochi rosii
o femele cu ochi rosii = 1/4
o femele cu ochi albi = 1/4
o masculi cu ochi rosii = 1/4
o masculi cu ochi albi = 1/4
- Concluzii:
v gena → culoarea alba a ochilor se exprima la ambele sexe
v gena → culoarea ochilor este localizata pe cromosomul X
- scoala lui Morgan → C.Bridges (1925) - teoria cromosomiala a ereditatii
- concluzie: legile lui Mendel = consecinta a repartitiei cromosomilor in meioza
o repartitia cromosomilor → distributia particulara a diferitilor loci →
o determinismul diferentelor fenotipice in ontogeneza
o legea I-a ← segregarea unei perechi de omologi in anafaza I din meioza
o legea a-II-a ← asortarea intamplatoare a omologilor in metafaza I
5. Fenomenul de linkage a genelor pe cromosom
descoperirea determinismului cromosomial al sexelor
o localizarea genelor pe cromosomii de sex → diferentierea sexuala
o a complicat genetica mendeliana
principiul segregarii ≠ pentru loci situati pe un acelasi cromosom
segregarea acestor alele - segregarea determinismului sexual
v gene sex-linkate (cuvantul englezesc linkage = inlantuite)
v trasaturi X-dominante sau X-recesive doar la ♀
v la ♂ - dominanta sau recesivitatea nu au sens - hemizigotie
v genele X-recesive mutante → boli hemofilia, daltonismul,
v genele Y-linkate - linie masculina → ereditatea holandrica
linkage genetic - W.Bateson, E.R.Saunders si R.Punnett (1905)
o P - mazare - linii pure
dublu homozigot dominante - fenotip flori purpurii - polen alungit
linii dublu homozigot recesive - fenotip flori rosii - polen rotund
o F1 - dublu heterozigoti - fenotipuri flori purpurii - polen alungit
o F2 raportul de asortare a fost:
74,6% (in loc de 56,25% = 9/16) flori purpurii - polen alungit
5,5% (in loc de 18,75% = 3/16) flori purpurii - polen rotund
5,5% (in loc de 18,75% = 3/16) flori rosii - polen alungit
14,4% (in loc de 6,25% = 1/16) flori rosii - polen rotund
? procentul de asortare a fenotipurilor (din P) - mai mare decat se asteptau
o frecventa ↑a polenului cu asocierea de gene din P
o asocieri de loci dominanti, respectiv recesivi ce pareau a fi cuplati
o explicatie: scoala lui Morgan experiente pe D. melanogaster
genele X-linkate - fenotip ochi albi-aripi miniaturale = cuplate
6. Linkage si recombinare genetica
recombinarea genetica - asocieri ale unor alele, care la genitori nu erau cuplate
o recombinarea genetica → fenotip recombinant
Morgan - recombinarea = schimb de material genetic intre omologi in meioza
Harriet Creighton si Barbara McKlintock (1931) - crossing-over
o recombinarea asociata cu schimb fizic de fragmente intre 2 omologi
7. Fenomenul de crossing-over si cartarea genelor
A.Sturtevant (1913) - a postulat ca:
o daca distanta dintre gene este determinata experimental
o se poate identifica si ordinea genelor pe cromosom
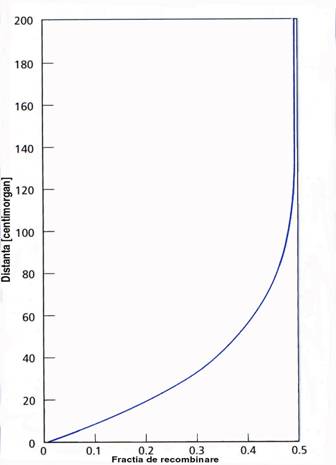
o frecventa crossing-over: distanta dintre loci - procentul de recombinare
o frecventa unui crossing-over este proportionala cu distanta dintre loci
o aplicatie: cartarea genica-distanta dintre gene = procente recombinare
o unitate de cartare genica - centimorganul [cM]
distanta dintre 2 loci la care frecventa de recombinare este 1% (recombinarea apare la 1% din procesele de crossing-over)
10 cM = 10 recombinari din 100 evenimente de crossing-over
grup de linkage = totalitatea genelor de pe un cromosom
o numar grupuri de linkage = numar cromosomi dintr-un set haploid
o determinarea localizarii unei gene - linii celulare hibride (hibridoame)
Figura 2-8. Relatia dintre fractia de recombinare si distanta dintre gene.
8. Conceptia clasica despre gene
8.1. Gena ca unitate de structura si functie
- unitatea structura: segment delimitat, continuu si indivizibil, dispus linear
- unitatea de functie: determina un anumit caracter fenotipic
pozitia genei pe un cromosom asociata cu un caracter fenotipic specific
genele se transmit de la o celula la alta prin cromosomi
8.2. Gena ca unitate de recombinare
profaza I - crossing-over = schimb reciproc de fragmente
cromosomiale
o genetica clasica - schimb de alele intregi
o secventa de alele cuprinde gene ce proveneau de la cei doi genitori
crossing-over = principalul mecanism de recombinare (diversificare) genetica
o responsabil de variabilitatea indivizilor unei specii →
o unitatea de recombinare
unitatea recombinare - segmental minim ce poate fi schimbat prin crossing-over
8.3. Gena ca unitate de mutatie
trasaturi avantajoase = caractere salbatece - codificate de alele salbatece
mutatiile → alela mutanta - codifica un caracter mutant
o fenotipic observabila - trasatura difera de cea salbateca
o duc la aparitia de variante noi ale unui caracter in populatiile umane
unitatea de mutatie: fragmentul cel mai mic care poate fi modificat
9. Modificarea paradigmei geneticii clasice
genetica clasica - nu rezolva problema naturii genei si a mecanismelor ereditatii
Beadle si Tatum (1941) - studiul caii metabolice a Arg pe Neurospora
o 3 tulpini distincte - necesitau mediu cu Arg pentru crestere
arg-1 arg-2 arg-3
↓ ↓ ↓
Enzima 1 Ezima 2 Enzima 3
Precursor → Ornitina → Citrulina → Arginina
rezultate:
o arg-1 creste in prezenta Orn si Cit - converteste precursorii in arginina
o arg-2 nu creste in prezenta Orn, dar creste in prezenta Cit
nu poate transforma Orn → Arg - poate converti Cit → Arg
o arg-3 nu creste in prezenta Orn si Cit - nu converteste substratele in Arg
concluzii:
o gena arg-1 → E-1 → transformarea precursori → Orn
o gena arg-2 → E-2 → transformarea Orn → Cit
o gena arg-3 → E-3 → transformarea Cit → Arg
ipoteza:
o reactiile biochimice decurg in etape
o fiecare etapa fiind catalizata de o enzima
o fiecare enzima este controlata de o gena specifica
prima piatra de hotar care a marcat drumul spre genetica moleculara
paradigama geneticii: o gena = un caracter → o gena = o proteina
proteinele - componente esentiale - nivelul celular de organizare al materiei vii