|
|
|
|
BIOLOGIE ANIMALA
CARACTERIZAREA GENERALA A ZONELOR UMEDE CERCETATE
1. Caracteristici hidrografice ale Raului Neajlov
Raul Neajlov, afluent pe dreapta tehnica a raului Arges, pe baza caracteristicilor de curgere este prezentat ca apartinand retelei hidrografice de tip dendritic a Romaniei, care caracterizeaza majoritatea raurilor din zona de campie si podis, diferentiate de celelalte retele prin asocierea ramurilor hidrografice sub forma de arbore. Desi situat in zona de campie, mai precis in Campia Burnasului raul Neajlov se numara printre raurile cu cele mai mari retele hidrografice (3 660 km2) ale tarii noastre. De asemenea, Neajlovul participa la formarea unuia dintre cele mai importante noduri de convergenta din zona de campie, la Stoenesti, la care conflueaza, alaturi de Neajlov raurile Calnistea, Dambovnic, Glavacioc. Coeficientul de sinuozitate caracteristic raurilor din zona de campie este cuprins intre 1,30 si 1,60. In cazul raului Neajlov, datorita diferentei mici de altitudine in profilul longitudinal al albiei coeficientul de sinuozitate se apropie de valoarea maxima insa lucrarile de amenajare realizate au avut ca rezultat scaderea nivelului de varsare a afluentilor , cresterea vitezei de curgere a apei in albie si reducerea gradului de sinuozitate in lunca Neajlovului. Dupa cum am mentionat anterior raul Neajlov se caracterizeaza printr-o suprafata de 3361 km2 a bazinului hidrografic, aflandu-se la o altitudine medie de 162 metri si un debit mediu al apei de 6,48 m3/s. In privinta valorilor scurgerii medii lunare se constata valori maxime in perioada de primavara (lunile martie-aprilie) si minime in perioada de vara-toamna (lunile august-septembrie).
Tipul de regim hidrografic (stabilit pe baza formei hidrografului, repartitiei sezoniere a scurgerii si surselor de alimentare)
In zona localitatii Comana raul Neajlov se incadreaza in Tipul "G" al scurgerii, in care scurgerea cea mai importanta este in primavara, urmand apoi, ca valoare, scurgerea din sezoanele de iarna si de vara. Alimentarea subterana variaza intre 10 si 30 % din scurgerea totala iar alimentarea superficiala este predominant nivala.
Traiectul initial al raului este descris in Tipul "F1", localizat in piemonturile sudice, unde scurgerea cea mai mare este in primavara, urmand apoi cea din iarna si vara. Alimentarea subterana reprezinta 20 - 40 % din scurgerea totala, iar alimentarea superficiala este mixta.
Scurgerea maxima: in majoritatea cazurilor debitele maxime anuale sunt de provenienta pluviala. In Campia Romana (zona localitatii Comana) aproximativ 70 % din debitele maxime au provenienta pluviala si numai 29 % au provenienta mixta. Analiza frecventei apelor mari si viiturilor realizata pentru posturile hidrometrice luate in considerare a aratat ca, in cursul anului, numarul maxim de varfuri are loc primavara (30 - 50 % din total), minimul fiind inregistrat toamna (10 - 20 %) sau iarna (5 - 30 %). Elementele caracteristice ale undelor de viitura singulare tip la posturi hidrometrice si volumele straturilor scurse la debitele maxime de asigurare (1%):
2. Caracterizarea padurii Comana
Studiul statiunii si al vegetatiei
Din punct de vedere geografic, teritoriul investigat este localizat in Campia Romana, districtul Campiei Burnasului (Burnasul estic), spre nord fiind delimitat de Campia Vlasiei, spre est de Campia Baraganului (Campia Mostistei), iar spre sud de lunca inundabila a Dunarii. Reteaua hidrografica este in general bine reprezentata, vaile raurilor fiind separate prin terase care dispar in zona de divagare a raurilor, cu interfluvii largi, terenul avand diferite inclinari si expozitii.
Ca forme depresionare de relief se intalnesc valcele, vai cu mlastini si numeroase microdepresiuni inchise, in care, functie de adancimea si intinderea lor s-au format soluri care corespund unor conditii de umiditate excesiva.
In zona studiata se intalnesc cele mai vechi forme de relief din campia Burnasului, campie tabulara cu terase.
Campia propriu-zisa (platforma Burnasului) are o altitudine cuprinsa intre 85 si 90 metri, prezentand diferente de nivel de aproximativ 30 metri in zona de nord a regiunii (spre raurile Neajlov si Arges) si mai mari, aproximativ 60 metri spre lunca Dunarii.
Terenul prezinta o inclinare slaba de la est spre vest si este strabatut de vai orientate de la sud spre nord, care se adancesc pana la 40 - 70 metri. In locurile plane apar depresiuni (podini), cu diametre cuprinse intre 500 si 1500 metri. Apa freatica se gaseste la aproximativ 30 metri. Adancimea mare a panzei de apa freatica face ca solurile di platforma Burnasului sa aiba caractere de soluri automorfe brun roscate, mai mult sau mai putin podzolite, cernoziomuri degradate.
Spre sud-est, aceasta platforma (a Burnasului) este marginita de o terasa inalta (terasa Greaca ), de 72-75 m altitudine absoluta, cu diferente de nivel de 56-58 m fata de lunca Dunarii. Apa freatica se gaseste la 30 m. Pe aceasta terasa nu se mai intalnesc paduri .
Campiile mai recente din Ocolul Silvic Comana sunt : Campia Vlascei 9campia Neajlovului), situata intre Arges si Calnistea, impartita in doua de Neajlov si Campul calnicului, situat la nord-est de Arges. Altitudinile relative sunt de aproximativ 30-40 m in Campia Neajlovului se de aproximativ 20m in Campul Calnoului. Apa freatica in aceste campii este de 10-20 m. Relieful lor este reprezentatat printr-o suprafata plana care coboara treptat spre su-est. ; se intalnesc depresiuni (crovuri) analoage cu cele din platforma Burnasului, de 2-6 m adancime, cu soluri pseudogleice podzolite de depresiune. Pe aceste campii mai recente nu s-a pastrat padurea, fiind inlocuite cu culturi agicole.
Luncile ocupa suprafete intinse in UP I si UP III ele prezinta diferentieri in functie de temperamentul raurilor si de materialele aduse.
Altitudinea variaza in jur de 80m, fiind cuprinsa intre 45m (in partea din aval a Argesului) si 60m (in partea de sud a ocolului).
Pantele sunt putin raspandite si apar ca forme de trecere intre unitatile geomorfologice (luncile raurilor si interfluvii)
Expozitia si inclinarea conditioneaza usoare modificari in regimul apei si in geneza solurilor.
Hidrologie
Suprafata paduroasa a O.S. Comana este strabatuta de raul Arges - afluent de ordinul I al Dunarii si de afluentii sai directi Neajlovul si Sabarul . debitul lor este permanent dar inconstant in timpul anului. In anii ploiosi se produc revarsari de ape cu insemnate pagube materiale.
In partea centrala a ocolului de-a lungul Neajlovului se afla balta Comana , care are un regim hidrologic variabil.
Influentele sistemului hidrologic asupra vegetatiei forestiere se resimt direct in zonele de lunca , prin frecventa inundatiilor, materialul aluvial depus, nivelul apei freatice care determina anumite tipuri de vegetatie caracteristice (zavoaie de plop si salcie, aninisuri, frasinete, sleauri de lunca) si influenta baltii Comana care creaza un microclimat mai umed. Reteaua hidrografica semipermeabila este constituita din vai secundare cu debit de apa numai in timpul ploilor si isi inceteaza activitatea, scurt timp dupa incetarea acestora. Aceste vai sunt scurte, destul de adanci, uneori cu versanti puternic inclinati si cu caracter torential.
Apa freatica , in campia tabulara, se afla la mari adancimi (30-40 m)
In sectoarele de lunca variaza intre 20 cm (in partile depresionate) si 2,5-3 m pe grinduri. Acest lucru influenteaza puternic atat repartitia speciilor forestiere cat si productivitatea si calitatea acestora. In lunca raului Arges in urma sapaturilor Canalului Dunare-Bucuresti apa freatica a coborat la 5-6 m, fapt care a condus la uscarea in masa a arboretelor de salcie, plop alb, plop negru si plop euramerican. In urma construirii canalului Dunare-Bucuresti, lunca Argesului a devenit inapta pentru speciile de zavoi (salcie, plop alb, plop negru) si plop euramerican ca urmare a coborarii nivelului apei freatice fapt care a dus la schimbarea statiunilor.
Neajlovul - rau imbatranit - prezinta meandre evoluate, un curs subsecvent si fenomene de divagare cu balti intinse (cum este cel de la Comana) Depresiunile acestui rau sunt aluviuni carbonatice fine, cu textura lutoasa sau luto-argiloase. Acesta este principala cauza care a dus la formarea de soluri si statiuni net diferite de cele de lunca Argesului. Vegetatia forestiera naturala este formata din frasinete, aninisuri, plopisuri de plop alb , sleauri de lunca.
Sabarul are importanta mai mica pentru sectorul silvic, prin faptul ca de-a lungul lui sunt putine paduri care prezinta caracteristici asemanatoare cu cele ale raului Neajlov. Materialele aluviale depuse sunt, deasemenea fine, fiind luto-argiloase, carbonatice.
O influenta asupra vegetatiei forestiere o au si baltile din lunca Neajlovului care creaza un microclimat mai umed in jur.
Geologie
Din punct de vedere geologic fata intregii campii Romane este acoperita de depozite de origine quaternara - loese si depozite mezozoice (cretacice, jurasice si triasice superioare) asezate peste cristalin.
In campia Burnasului, peste fundamentul prebalcanic care cuprinde formatiuni geologice mai vechi decat Cretacicul intalnit la aproximativ 100m adancime si peste depozitele pliocene de deasupra acestui nivel, formate din argile, nisipuri si pietrisuri, apare quaternarul reprezentat in partea sa inferioara prin pietrisurile de Fratesti. Aceste pietisuri care se intalnesc in straturi de 1-17 m grosime apar pe la 40-45 m adancime si sunt alcatuite din elemente de gnaise, porfire, corneene, silex, cuart filonian, gresii varuconice si glauconitice, marme calcaroase si calcare dretaice, riolite etc. de provenienta nu numai carpatica, ci si balcanica.
Peste aceste pietrisuri apar depozite marnoase, in mare parte erodate, nisipuri marnoase sau marne nisipoase, care dezvoltate in general pe aproximativ 6 m sunt adesea o cauza a declansarii de alunecari de teren. Peste aceste depozite se intalnesc materiale fine eoliene cu cuverura luto-argiloasa si loesolide , dezvoltate pe 10-20m, datand cu incepere din Riss si pana in Holocen, se disting in afara solului actual, cel putin doua orizonturi clare de soluri fosile. Depozitele luto-argiloase din apropierea suprafetei formeaza cel mai adesea materialul parental al solurilor de pe campia inalta a plaformei Burnasului.
Depozitele de cuvertura din Campia Neajlovului si Campia Calnaului sunt mai putin adanci ca in platforma Burnasului, dar prezinta si ele doua soluri fosile.
Pe bordura Campiei Neajlovului si Campia Calnaului se dezvolta terasele pleistomene. Terasa superioara, cu o altitudine relativa de 20-22 m are apa freatica la 15-20 m adancime, terasa medie, cu altitudine relativa de 12-16 m are apa freatica in 8-18 m adancime. Atat pe terasa superioara cat si pe cea medie, se dezvolta soluri auto-morfe pe luturi argiloase eoliene care permit datarea lor cu wurn 1 si wurn 2. Terasa inferioara, aflata la altitudini relative 5-7 m, are depozite de cuvertura mai subtire si mai putin argiloase, de o varsta mai tanara (materialul depus in wurn 3 si terasa morfologica sapata in Halocen). Apa freatica este la 5-10 m. Solurile acestor terase sunt foarte variate, in functie de substrat, pe materiale mai nisipoase soluri brune roscate, iar pe cele argiloase cernoziomuri freatic umede. In depresiunile acestei terase exista pe alocuri soluri cu procese de solonetizare.
Luncile formate numai din materiale halocene (uneori foarte recente), prezinta diferente in functie de temperamentul raurilor si de materialele depuse. Argesul - un rau tanar - aduce aluviuni predominant mai nisipoase, de-a lungul Neajlovului si Calnistei predomina aluviunile mai argiloase, in care procesele de argilizare sunt mult mai frcvente
Se observa o diferentiere si in profitul transversal al luncilor. In apropierea talvegului sunt aluviunile cele mai recente si cele mai grosiere (in general nisipoase), todeauna nesolificate. Inundabilitatea in aceste sectoare este frecventa si apa freatica foarte aproape de suprafata. Urmeaza, de obicei, un grind de aluviuni nisipoase, cu apa freatica la adancime mai mare (uneori peste 1,5 m); pe acest grind se intalnesc obisnuit soluri aluviale mai putin gleizate decat restul luncii. Dupa acest grind apare lunca cu cele mai argiloase sedimente, cu cea mai accentuata gleizare si cu apa freatica la mai putin de 1m adancime. Aici s-au dezvoltat soluri cu caractere hidromorfe accentuate. Sporadic se intalnesc portiuni de lunca drenata mai intens, cum este aceea de langa Campurile (U.P.III), unde se dezvolta soluri mai evaluate, de tipul solului brun sub stejarete-sleauri, cu bogata flora de mull.
La confluenta Neajlovului si Sabarului cu Argesul, datorita nivelului freatic ridicat si incetinirii vitezei de scurgere a apei, evapotranspiratia este mai accentuata, ceea ce determina acumularea de saruri solubile (Gradistea UP III)
Climatologie
Ca urmare a amplasarii lor, padurile O.S. Comana se afla in plin climat continental de campie, caracterizat prin veri foarte calde si ierni foarte reci.
Dupa raionarea climatica di « Monografia geografica », teritorial considerat apartine formulei climatice cfax, care ocupa partea sud-vestica si formulei Dfax, care acopera restul ocolului. Incadrarea padurilor din cuprinsul ocolului silvic Comana in cele doua formule climatice se traspune in vegetatie prin existenta bine manifestata a doua nivele de vegetatie mult diferite ; asociatii de paduri mezofile- caracteristice zonei forestiere-situate la nord, si asociatii vegetale cracteristice zonei de silvostepa in sud. Intre acestea exista o gama larga de tranzitii, cu caracter zonal, determinate de modificari in climat si in sol.. Pe langa factorii mentionati mai survin unele modificari ale acestei zonalitati, determinate de unii factori locali, azonali, care se reflecta in caracterele fizice sau fizico-climatice ale solurilor (compacitatea, umiditatea etc)
Urmarind seria succesiunilor zonale de la nord la sud, in cuprinsul ocolului se pot separa urmatoarele subzone fito-climatice ;
subzona padurilor de stejar pedunculat, care ocupa o suprafata destul de restransa si se caracterizeaza prin ominarea in arborete a stejarului pedunculat, insotit de numeroase elemente de sleau si cu flora erbacee formata din specii de mull ;
subzona padurilor de tranzitie de stejar pedunculat si cereto-garnitete, care ocupa o fasie mai extinsa si cuprinde arborete in care quercineele mezo si termofile, insotita uneori si de elemente de sleau ;
subzona padurilor termofile - cea mai bine reprezentata ca suprafata - care este caracterizata din paduri de cer si garnita, cu flora caracteristica.
Aceasta subzona face tranzitia intre zona forestiera si cea a silvostepei.
zona silvostepei - pe o suprafata destul de redusa in extremitatea sud vestica a ocolului - cu paduri de stejar brumariu in amestec cu alte quercinee semixerofite si xerofite (cer, garnita, stejar pufos) si cu flora erbacee in covor continuu, formata din specii xerofile si semixerofile.
In afara acestei zonalitati bioclimatice, mai sunt prezentate o serie de formatii vegetale interzonale - de lunca - a caror diversitate in spatiu este determinata de natura materialului aluvial (textura) si regimul apei din sol, caldura si umiditatea atmosferica nu influenteaza aceste formatii vegetale. Indicel de ariditate de Martonne este de 24-28 (valoarea mai mare corespunde padurilor de sleau cu stejar pedunculat)
Regimul termic
Temperatura medie anuala prezinta variatii foarte mici, in cadrul ocolului, intre 10,80C si 11,2 0C. Temperatura medie a lunii ianuarie este de aproximativ -30C. a lunii ianuarie este de mare importanta, intrucat se separa in sistemul Koppen, climatele temperate (c.f.) de cele boreale (z.f.) Aceasta izoterma trece prin mijlocul ocolului ; partea sudica a teritoriului are o temperatura medie a lunii ianuarie putin mai ridicata decat -30C. temperatura medie a lunii iulie este de aproximativ 250C, fara variatii semnificative. Temperatura minima absoluta inregistrata a fost de -31,20C la 24.I.. 1942, iar temperatura maxima absoluta inregistrata a fost de 40,30C la 29.VII.1928.
Prima zi de inghet apare in jurul datei de 1.XI, iar ultimul inghet in jurul datei de 11.IV. Nu s-au constatat geruri tarzii sau timpurii care sa fi avut influenta asupra vegetatiei forestiere. Se poate trage concluzia ca perioada de vegetatie este de destul de lunga si regimul termic este favorabil vegetatiei forestiere.
Regimul pluviomertic
Precipitatiile medii anuale, care cad in raza O.S. Comana ating in jur de 560 mm in partea de nord a ocolului si de 520 mm in centrul masivului padurea de la sud de Comana. Precipitatiile sunt repartizate in tot cursul anului cu oarecare accentuare la inceputul verii (suma medie a precipitatiilor lunii iunie ajung la 76 mm).
O diminuare a cantitatilor se remarca la inceputul toamnei si in timpul iernii (cu deosebire in luna februarie).
Repartizarea pe anotimpuri se prezinta astfel : iarna 76-100mm, primavara 125-150 mm, vara 150-175 mm, toamna 100-125 mm.
Indicele de ariditate anual este de 24-26 (valoarea mai mare corespunzand padurilor de sleau cu stejar pedunculat).
Evapotranspiratia potentiala anuala, in jur de 730 mm, valoare care depaseste cu mult cantitatea precipitatiilor anuale. Aceasta depasire provine din depasiriel lunare din perioada de vegetatie (aprilie- octombrie) valoarea maxima medie lunara a evapotranspiratiei potentiale se realizeaza in luna iulie. Evapotranspiratia reala (305 mm) este inferioara evapotranspiratiei potentiale si chiar a sumei precipitatiilor. Deficitul in ceea ce priveste rezervele de apa, vegetatia se bazeaza mai mult pe rezervele din sol, decat pe apa din precipitatii.
Deficitul de umiditate deteremina in principal repartitia zonala a vegetatiei forestiere in ocolul Comana.
Regim eolian
Teritoriul in care sunt amplasate padurile din O.S. Comana este expus in intregime actiunii vantului.
Vanturile predominante sunt cele din nord-est si sud-vest, cu intensitatea cea mai mare iarna, atingand dupa scara Beaufort gradul 5-7, ceea ce corespunde cu o viteza de 27-54 Km/ora.
Intensitatea maxima a acestor vanturi se produce pe o durata medie de pana la 10 zile anual, in restul timpului intensitatea fiind mai scazuta.
Vanturile au o influenta daunatoare asupra vegetatiei forestiere, in special cele uscate si foarte calde din timpul verii, prin scaderea umiditatii din aer si sol, marirea evapotranspiratiei. Doboraturi de vant nu se produc datorita faptului ca speciile forestiere din cadrul ocolului sunt rezistente la actiunea vantului, iar arboretele sunt in marea lor majoritate tinere.
Concluzii privind conditiile climatice
Luand in considerare valorile Indicilor de ariditate Martonne, care exprima ariditatea regiunii prin raportul dintre precipitatii si temperatura se pot face precizari necesare privind conditiile climatice ale teritoriului in care se afla situat O.S. Comana.
In perioadele de vegetatie indicii de ariditate coboara sub 24, ajungand chiar pana la 18, ceea ce caracterizeaza conditii de silvostepa.
Conditiile pentru vegetatia forestiera rezulta si din valoarea deficitului de precipitatii, comparativ cu valoarea evapotranspiratiei potentiale, care se prezinta la aproximativ 200 mm, deci o cantitate apreciabila fata de valoarea precipitatiilor anuale.
Pedologie
Conditiile climatice, formele de relief si materialul parental au determinat formarea de tipuri si subtipuri genetice de sol, caracteristice regiunii (brune roscate de padure in diverse stadii de podzolire).
De-a lungul raurilor (Arges, Neajlov, Calnistea) sunt soluri aluviale, cu texturi diferite, functie de materialele aluviale depuse si care in raport cu frecventa si durata inundatiilor prezinta diferite stadii de evolutie spre tipul normal de sol.
S-au identificat soluri brune roscate (29 % din suprafata) in UP I - VII, soluri brune roscate luvice (37 % in UP III, IV,V,VI,VIII), brune eumezobazice (9 % in UP I, III), cernozion cambic (10 % in UP V, VII, VIII) in campie medie si soluri aluviale (12%), in lunca Argesului si Neajlovului in unitatile de productie I si III.
studiul Vegetatiei caracteristice regiunii
1. Cadrul natural
Din punct de vedere geobotanic, sectorul care face obiectul prezentului proiect se afla amplasat (dupa I. Dragu si colab., 1999) in zona padurilor de foioase (nemorala), subzona silvostepei (fig. 1).
Vegetatia forestiera este formata din paduri de Quercus frainetto, de Quercus cerris si de amestec al acestor doua specii, fiind dezvoltata pe soluri brune roscate luvice cu textura luto - argiloasa; in luminisuri si raristi se intalneste frecvent vegetatie ierboasa de stepa in care domina Festuca valesiaca. Mentionam de asemenea existenta in padurea Comana a doua specii rare: Paeonia peregrina si Ruscus aculeatus.
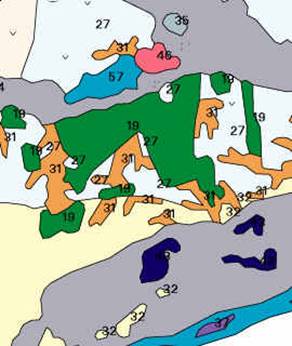
Fig. 1. Harta geobotanica a sectorului studiat (extras din Harta Geobotanica a Romaniei sc. 1:500.000)
LEGENDA
19. Paduri de Quercus frainetto, Quercus cerris etc.
27. Culturi in locul vegetatiei de silvostepa
31. Vegetatie de stepa pe coaste cu Botriochloa ischaemum, Festuca valesiaca etc.
32. Culturi in locul vegetatiei de antestepa
35. Paduri de Quercus robur + Fraxinus angustifolia, Ulmus minor etc.
37. Zavoaie de Populus sp., Salix sp., + Alnus glutinosa, Quercus robur, Fraxinus angustifolia etc. (in lunca Dunarii)
43. Vegetatie cu asociatii higrofile de Phragmites australis, de Typha angustifolia etc.
45. Pajisti cu asociatii de Puccinelia distans, de Camphorosma annua etc.
57. Vegetatie de balti si lacuri Vegetatia de balti si lacuri este formata din asociatii higrofile de Phragmites australis, de Typha angustifolia, de Schoenoplectus lacustris, de Glyceria maxima etc., in complex cu asociatii natante si submerse de Myriophyllum sp., de Ceratophyllum sp., de Nymphaea alba, de Nymphoides peltata, de Hydrocharis morsus-ranae, de Trapa natans, de Stratiotes aloides, de Nuphar luteum, de Lemna sp., de Potamogeton sp., alge etc.
2. Metoda folosita
Lucrarile de teren au constat in efectuarea de relevee geobotanice in 10 suprafete pentru monitorizarea vegetatiei hidrofile si o suprafata preliminara in zona malului acoperita cu vegetatie forestiera, constand in:
instalarea unor suprafete volante in care s-au realizat conspecte floristice totale;
recoltarea de material botanic floristic pentru confirmarea sau identificarea in laborator a incadrarii taxonomice a speciilor.
S-a folosit metoda de inventariere globala pe o suprafata de 100 m2, estimarea gradului de acoperire a speciilor fiind procentuala.
Raporturile cantitative si calitative au fost stabilite pe baza indicatorilor fitocenologici: abundenta + dominanta si constanta dupa metoda Braun-Blanquet, exprimate procentual.
In laborator s-a facut verificarea incadrarii taxonomice a speciilor de plante dupa Flora Romaniei vol. I-XIII. Nomenclatura utilizata este conforma cu Flora Europaea (Tutin et al., 1968-1980; Tutin et al., 1993) pentru plantele vasculare.
Lucrarile de birou au constat in prelucrarea si interpretarea datelor de teren si laborator.
Rolul fitocenotic al componentilor asociatiilor vegetale a fost apreciat atat dupa speciile dominant-edificatoare, care reflecta in cel mai inalt grad conditiile ecologice asupra caruia actioneaza, cat si dupa speciile caracteristice indicatoare, care oglindesc fidel actiunea anumitor factori ecologici si istorici. S-a acordat atentie si speciilor insotitoare, care, desi putine cantitativ, intra in compozitia floristica a asociatiei. Fiecare asociatie a fost analizata si prin prisma bioformelor si a elementelor floristice (Beldie, 1977, 1979).
Incadrarea bioclimatica este conforma cu Harta geobotanica a Romaniei (Dragu si colab., 1989).
Denumirea asociatiilor vegetale a fost facuta atat dupa metoda speciilor dominante, cat si dupa metoda fitocenologica (clasificarea asociatiilor si incadrarea fitocenologica s-a facut dupa Sanda si colab., 2001).
3. Rezultate obtinute
Releveele geobotanice cuprind un total de 41 de speciile de plante care fac parte din 24 familii de plante superioare si una de alge (tabelul 1).
In zona lacului Comana, majoritatea speciilor identificate sunt plante ierboase hidrofite, terestre sau natante, frecvent submerse (Myriophyllum verticillatum) cu centrul de raspandire circumpolar sau cosmopolit, mai rar eurasiatic (Hydrocharis morsus-ranae, Sparganium erectum) sau european (Mentha aquatica, Polygonum mite); la noi in tara aceste specii pot fi intalnite frecvent in baltile si mlastinile din stepa si pana in etajul padurilor de fag montan.
Spectrul ecologic evidentiaza speciile adaptate conditiilor de umiditate (ultrahigrofite si higrofite) specifice malurilor (Typha sp.) si apelor stagnante sau incet curgatoare (Myriophyllum verticillatum, Lemna minor etc.), putin adanci (sub 2 m), cu cerinte mari privind aprovizionarea cu substante nutritive (eutrofe - mezotrofe), uneori chiar nitrofile (Polygonum mite, Urtica dioica). Se intalnesc de asemenea specii indicatoare ale lipsei carbonatilor (Hydrocharis morsus-ranae), ale unui pH acid (Lemna minor) sau moderat acid (Lythrum salicaria) si uneori ale prezentei humusului hidromorf (Lythrum salicaria, Mentha aquatica). Au fost remarcate de asemenea si specii indicatoare ale unei slabe tendinte de mineralizare (Berula erecta - care este insa si o specie toxica).
In zona malurilor, acoperite cu vegetatie forestiera, rezultatele preliminare de anul acesta remarca faciesul cu Fraxinus excelsior, insotit frecvent de Ulmus minor, Acer campestre, Cornus sanguinea, Prunus spinosa, Crataegus monogyna, Rubus hirtus etc. Dintre speciile ierboase se remarca: Agrimonia eupatoria, Carex sylvatica, Urtica dioica, Lysimachia nummularia etc. Fitocenozele au un grad mediu de incheiere (acoperirea stratului arborescent este de 60%) fapt care a favorizat dezvoltarea arbustilor (grad de acoperire de 45%) a si ierburilor (grad de acoperire 75%).
Din punct de vedere fitocenologic au fost identificate 4 asociatii vegetale care se grupeaza astfel:
I. Vegetatia acvatica (Aquiherbosa)
A. Clasa Lemnetea O. de Bolós et Masclans 1955 (vegetatia acvatica nefixata)
a. Ordinul Lemnetalia minoris O. de Bolós et Masclans 1955 (vegetatia plutitoare)
i. Alianta Lemnion minoris O. de Bolós et Masclans 1955 (grupari de lintita)
1. Asociatia Lemnetum minoris Soó 1927
B. Clasa Potamogetonetea pectinati R.Tx et Prsg. 1942 (vegetatia acvatica fixata)
a. Ordinul Potamogetonetalia pectinati W. Koch 1926 (vegetatia de apa dulce)
i. Alianta Potamogetion lucentis Rivas Martinez 1973 (grupari submerse + natante)
2. Asociatia Myriophyllo - Potametum lucentis Soó 1934
subasociatia myriophylletosum verticillati
II. Vegetatia palustra (Paludes)
A. Clasa Phragmitetea australis R.Tx. et Prsg. 1942 (vegetatia mlastinilor eutrofe)
a. Ordinul Phragmitetalia Koch 1926 (mlastini cu ierburi inalte)
i. Alianta Phragmition communis Koch 1926 (trestiiso - papurisuri)
3. Asociatia Scirpo-Phragmitetum W. Koch 1926
subasociatia phragmitetosum
subasociatia typhosum latifoliae
III. Vegetatia lemnoasa (Lignosa)
A. Clasa Querceto - Fagetea Br.-Bl. et Vlieger in Vlieger 1937 em. Borhidi 1996 (paduri de foioase mezofile)
a. Ordinul Fraxinetalia Scam. et Pass. 1959 (paduri de lunca cu esente tari)
i. Alianta Ulmion Oberd. 1953 (stejarisuri cu frasin si ulm)
4. Asociatia Querceto-Fraxineto-Ulmetum Issler 1924
VEGETATIA PALUSTRA: Studiul geobotanic a pus in evidenta predominarea stufarisurilor, grupate din punct de vedere fitocenologic in clasa Phragmitetea australis R.Tx. et Prsg. 1942, clasa care inglobeaza vegetatia dominata de trestiiso-papurisuri. Cenozele acestei clase se instaleaza incepand dinspre mal sau, fiind favorizate de nivelul mic al lacului, in zonele colmatate din mijloc. Asociatia se incadreaza in ordinul Phragmitetalia Koch 1926 care reuneste mai ales fitocenozele higrofile din zona de campie, sarace in specii, situate la marginea apelor lente sau stagnante si in alianta Phragmition communis Koch 1926. Speciile caracteristice aliantei sunt principalii edificatori ai vegetatiei palustre: Phragmites australis, Typha latifolia, T. angustifolia etc.
asociatia de Phragmites australis (Scirpo-Phragmitetum W. Koch 1926) este asociatia cu cea mai mare amplitudine datorita colmatarii lacului; principalele specii de recunoastere ale asociatiei formeaza pe alocuri subasociatii, dupa cum unele dintre ele capata rol dominant si edificator; in zona de mal si in arealele colmatate predomina Phragmites australis dupa care urmeaza o fasie relativ ingusta cu Typha latifolia, local si Typha angustifolia. Au fost separate astfel doua subasociatii: phragmitetosum si typhosum latifoliae (tabelul nr.2). In compozitia asociatiei a fost identificat un numar foarte mic de specii, majoritatea fiind specifice clasei Phragmitetea.
Spectrul biologic scoate in evidenta predominarea cvasi-totala a hidrofitelor. Hemicriptofitele se intalnesc sporadic, in papuris, cu precadere in zona adiacenta malurilor (langa ponton - releveu nr. 4), majoritatea fiind specii higrofite - ultrahigrofite.
In asociatia de trestie cu papura, elementele cosmopolite si circumpolare sunt mai bine reprezentate, celelalte elemente (european si eurasiatic) avand aparitii sporadice.
Importanta economica a trestiiso-papurisurilor este redusa, ele fiind daunatoare pisciculturii, indiferent daca sunt situate la marginea sau in centrul lacurilor.
VEGETATIA ACVATICA NATANTA: Vegetatia acvatica plutitoare este formata in majoritate din fitocenozele de lintita (Lemna minor). Din punct de vedere fitocenologic aceste cenoze sunt incadrate in clasa Lemnetea O. de Bolós et Masclans 1955, ordinul Lemnetalia minoris O. de Bolós et Masclans 1955, alianta Lemnion minoris O. de Bolós et Masclans 1955.
asociatia de Lemna minor (Lemnetum minoris Soó 1927) populeaza zonele cu ape linistite, adesea la adapostul vegetatiei palustre, sau cu slabi curenti de fund si suprafata din mijlocul lacurilor. Este o asociatie in general monodominanta in care patrund sporadic indivizi razleti ai altor specii acvatice si palustre (Spirodela polyrhiza, Sparganium erectum, Hydrocharis morsus-ranae, Lythrum salicaria etc.), cu precadere in situatiile in care asociatia patrunde printre indivizii de trestie si papura (Spirodela polyrhiza poate suporta un anumit grad de umbrire).
Dintre bioforme predomina hidrofitele, iar in ceea ce priveste caracterul fitogeografic al asociatiei, el este dominat de elementele cosmopolite, elementele circumpolare si eurasiatice ocupand un loc secundar.
Importanta economica a asociatiile de lintita este redusa, ele fiind daunatoare pisciculturii, deoarece stratul compact de lintite impiedica patrunderea razelor calorice in adancimea apelor; pe langa faptul ca apele de sub stratul de lintite sunt reci, ele sunt sarace si in oxigen iar prin degajarea de H2S rezultat din descompunerea substantelor organice provoaca intoxicarea pestilor.
VEGETATIA ACVATICA SUBMERSA: Vegetatia acvatica fixata prin sistemul radicular de substratul aluvial este in mare parte dominata de asociatia submersa in care elementul dominant edificator il reprezinta Myriophyllum verticillatum.
asociatia de Myriophyllum verticillatum (penita) este considerata din punct de vedere fitocenologic o subasociatie (myriophylletosum verticillati Soó 1957) a asociatiei Myriophyllo - Potametum lucentis Soó 1934, asociatie frecventa in apele statatoare sau slab curgatoare, cu adancimi intre 0,8-1,5 m. Acest tip de fitocenoze sunt incadrate in clasa Potamogetonetea pectinati R.Tx et Prsg. 1942 (fitocenoze acvatice submerse sau emerse din zona de campie) ordinul Potamogetonetalia pectinati W. Koch 1926 (fitocenozele acvatice fixate), alianta Potamogetion lucentis Rivas Martinez 1973.
Dintre bioforme predomina hidrofitele, iar in ceea ce priveste caracterul fitogeografic al asociatiei, el este dominat de elementele circumpolare si cosmopolite, elementele eurasiatice ocupand un loc secundar.
Aceasta asociatie reprezinta o etapa avansata a evolutiei fitocenotice a vegetatiei acvatice fata de cea anterioara. Ea are un rol important in procesele de purificare a apei, reprezentand de asemenea un biotop esential al faunei de nevertebrate (caracteristica a ordinului).
Incadrarea fitocenologica a asociatiilor vegetale acvatice sau palustre a intampinat numeroase dificultati datorate pe de o parte omogenitatii floristice, cu putine specii caracteristice si diferentiale, iar, pe de alta parte, integrarii cenotice reduse si autonomiei mai mari a sinuziilor pe masura ce adancimea apei este mai mare, ce a condus uneori la separarea lor in asociatii distincte (asociatii de Myriophyllum verticillatum si asociatii de Lemna minor). De asemenea, unele cenoze apartinand la unitati distincte (cum sunt cele trei asociatii descrise anterior) se pot intrepatrunde, integrandu-se in cenoze complexe.
VEGETATIA FORESTIERA : Rezultatele preliminare an remarca pe malurile lacurilor, cu soluri gleizate, cu apa freatica la adancime mica, un facies al asociatiei de Quercus robur cu Fraxinus excelsior si Ulmus minor in care elementul dominant edificator il reprezinta frasinul. Din punct de vedere fitocenologic asociatia se incadreaza in clasa Querco - Fagetalia Br.-Bl. et Vlieg. 1937, care inglobeaza padurile de foioase mezofile, de pe soluri eutrofe si mezotrofe. Ordinului Fraxinetalia Scam et Pass. 1959 ii sunt caracteristice padurile de lunca constituite din stejar, ulm si frasin, in care, de multe ori, ulmul sau frasinul (ca in cazul de fata) ating proportia de facies. Primavara, solul acestor paduri este foarte umed pana la mlastinos, iar vara este uscat; apa freatica se afla aproape de suprafata solului. Paduri asemanatoare din silvostepa sudica au fost denumite de S. Pascovschi si V. Leandru (1958) sleau normal de lunca din regiunea de campie.
- faciesul cu Fraxinus excelsior al asociatiei Querceto-Fraxineto-Ulmetum Issler 1924, este intalnit pe malul lacurilor, fiind invecinat in zona teraselor cu padurile de cer si garnita, de unde patrund uneori elemente subtermofile (Polygonatum latifolium).
Spectrul biologic evidentiaza predominarea exclusiva a fanerofitelor in straturile arborescent si arbustiv, si a hemicriptofitelor - local chamefite (Lysimachia nummularia) si geofite (Polygonatum latifolium) - in stratul erbaceu.
Spectrul ecologic evidentiaza speciile eutrofe - mezotrofe, mezoxerofite - mezohigrofite, local cu regim hidric alternant (Lysimachia nummularia, Carum carvi etc.). Sunt specii frecvent intalnite pe soluri eu-mezobazice, moderat acide - slab alcaline, bogate in humus de tip mull, afanate, cu textura mijlocie - usoara, gleizate sau temporar submerse. Local se intalnesc specii preferential calcicole si nitrofile.
In linii mari fitocenozele pastreaza caracterul asociatiilor europene, primind in stratul erbaceu numeroase specii eurasiatice, sub-mediteraneene (Carex sylvatica, Primula veris), local circumpolare (Geum urbanum) sau pontice (Polygonatum latifolium).
Desi reduse ca suprafata, padurile de lunca din aceasta zona prezinta o deosebit de mare importanta ca moderator al microclimatului arid din zona de campie.
Incadrarea fitocenologica a asociatiilor vegetale forestiere este preliminara, ea urmand a fi definitivata anul viitor prin efectuarea de relevee geobotanice atat in zona de lunca cat si pe campul inalt.
4. Succesiunea vegetatiei
Rezultatele preliminare obtinute in acest an ne ofera unele indicatii privind succesiunea probabila a vegetatiei in zona lacului Comana. Astfel, primele hidrofite care populeaza suprafata apelor sunt speciile plutitoare, nefixate de substrat prin radacini, formand fitocenozele natante (asociatiile de lintite).
Cu timpul, fitocenozele natante cedeaza locul hidrofitelor fixate de substrat prin radacini (asociatiile de penita) si raman dispersate sau se retrag spre tarmurile lacurilor sau ale canalelor. Substantele organice rezultate in urma descompunerii hidrofitelor, depozitate pe fundul apelor, favorizeaza dezvoltarea vegetatiei submerse si natante fixate de substrat.
Prin acumularea continua a substantelor organice, procesul de colmatare se accelereaza, creand conditii favorabile vegetatiei emerse palustre cu trestiis si papuris.
De la vegetatia palustra, in urma uscarii solului se face trecerea catre sleaurile de lunca, sau, pe terenurile mai ridicate, acestea evolueaza spre cerete (probabil prin asociatii de tranzitie de tipul Lythro - Calamagrostetum, Populeto - Salicetum, Pruno - Crataegetum):
Plante superioare (macrofite) inventariate in Balta Comana
1. Dryopteris thelipteris (L) A. Gray
2. Marsilea quadrifolia L.
3. Azola filiculoides Lam.
4. Salvinia natans (L.) All.
5. Polygonum amphibium L.
6. Polygonum amphibium L.f. aquaticum Leyss
7. Rumex aquaticus L.
8. Myosoton aquaticum L. Moench.
9. Saponaria officinalis L.
10. Batrachium aquatile (L.) Dumort.
11. Batrachium aquatile (L.) Dumort f. heliophylus Beck.
12. Batrachium divaricum (Schrank) Schur.
13. Batrachium trichophyllum (Chaix.) F. Schultz
14. Caltha palustris L.
15. Nuphar luteum L.
16. Nymphaea alba L.
17. Ceratophyllum demersum L.
18. Ceratophyllum submersum L.
19. Rorripa amphibia (L.) Bess.
20. Elatine alsinastrum L.
21. Myriophillum spicatum L.
22. Hippuris vulgaris L.
23. Oenanthe aquatica (L.) Poiret
24. Menyanthes trifoliata L.
25. Callitriche verna L.
26. Mentha aquatica L.
27. Veronica anagallis-aquatica L.
28. Veronica beccabunga L.
29. Utricularia vulgaris L.
30. Potamogeton crispus L.
31. Potamogeton lucens L.
32. Potamogeton natans L.
33. Potamogeton pectinatus L.
34. Najas minor All.
35. Iris pseudacorus L.
36. Glyceria maxima (Hartm.) Holmberg
37. Glyceria fluitans (L.) R. Br.
38. Phragmites australis (Cav.) Steudel
39. Lemna gibba L.
40. Lemna minor L.
41. Lemna trisulca L.
42. Sparganium ramosum Huds.
43. Typha angustifolia L.
44. Typha latifolia L.
45. Heleochais palustris (L.) R. Br.
46. Schoenoplectus lacustris (L.) Palla.
47. Schoenoplectus tabernaemontani (C.C. Gmelin) Palla.
Specii de alge inventariate in Balta Comana
1. Anabaena variabilis Kutz.
2. Oscillatoria brevis (Kutz.) Born.
3. Lyngbia martensiana Menegh.
4. Vaucheria dichotoma (L.) Ag.
5. Fragilaria virescens Raefs.
6. Synedra acus Kutz
7. Synedra rumpens Kutz
8. Synedra vaucheriae Kutz
9. Cocconeis placentula Ehrh.
10. Navicula cincta (Her.) Kutz
11. Navicula kryptocephala Kutz
12. Pinnularia viridis (Nitzach.) Ehr.
13. Caloneis amphisbaena (Bory.) Cl.
14. Gyrosigma acuminatum (Kutz) Rabenh.
15. Gyrosigma peisona (Grun.) Hust.
16. Pleurosigma angulatum (Quek.) W. Sm.
17. Cymbella amphicephala Nag.
18. Cymbella cuspidata Kutz.
19. Nitzschia acuminata (W.SM.) Grun.
20. Nitzschia apiculata (Greg.) Grun.
21. Nitzschia linearis (Ag.) W. Sm.
22. Nitzschia stagnorum Rabenh.
23. Surirella ovalis Breb.
24. Campylodiscus noricus Her.
25. Scenedesmus quadricauda (Turp.) Breb.
26. Ulotrix zonata (Web. Et Mohr.) Kutz
27. Enteromorpha percusa Ag.
28. Cladophora glomerata (L.) Kutz
29. Oedogonium sp.
30. Spyrogyra sp.
31. Chara vulgaris L.
32. Euglena ehrenbergii Kleis
33. Euglena polymorpha Dang.
34. Phacus pyrum (Her.) Stein.
Specii de plante inventariate in Padurea Comana
I. Rezervatia cu Ruscus aculeatus
1. Corylus avellana L.
2. Carpinus betulus L.
3. Quercus cerris L.
4. Quercus frainetto Ten.
5. Quercus pedunculiflora C. Koch.
6. Quercus robur L.
7. Ulmus foliacea Gilib.
8. Fraxinus excelsior L.
9. Fraxinus ornus L.
10. Ligustrum vulgare L.
11. Cornus sanguinea L.
12. Cornus mas L.
13. Evonymus verrucosa Scop.
14. Evonymus europaea L.
15. Staphylea pinnata L.
16. Tilia tomentosa Moench
17. Acer campestre L.
18. Acer tataricum L.
19. Acer platanoides L.
20. Sorbus torminalis (L.) Cr.
21. Pirus piraster (L.) Med.
22. Malus silvester (L.) Mill.
23. Crategus monogyna Jacq.
24. Holosteum umbellatum L.
25. Moehringia trinervia L. Clairv.
26. Anemone ranunculoides L.
27. Ficaria verna Huds.
28. Helleborus odorus W. et. K.
29. Asarum europaeum L.
30. Hypericum hirsutum L.
31. Corydalis cava (L.) Scweigg. Et. Koerte
32. Corydalis cava ssp. marchalliana (Pall.) Hay.
33. Corydalis solida (L.) Sn.
34. Alliaria petiolata (Bieb.) Cavara et Grande
35. Dentaria bulbifera L.
36. Geum urbanum L.
37. Fragaria vesca L.
38. Astragalus glycyphillos L.
39. Lathyrus niger (L.) Berh.
40. Lathyrus vernus (L.) Bernh.
41. Trifolium medium L.
42. Geranium robertianum L.
43. Euphorbia amygdaloides L.
44. Mercurialis perennis L.
45. Dictamnus albus L.
46. Viola canina L.
47. Viola sylvestris Lam.
48. Sanicula europaea L.
49. Vincetoxicum officinale Moench
50. Asperula taurina L.
51. Asperula odorata
52. Lithospermum purpureo-coeruleum L.
53. Pulmonaria mollissima Kern.
54. Pulmonaria officinallis L.
55. Lamium maculatum L.
56. Galeopsis speciosa Mill.
57. Melissa officinalis L.
58. Salvia glutinosa L.
59. Valeriana officinalis L.
60. Campanula macrostachya W. et. K.
61. Campanula ranunculoides L.
62. Campanula sibirica L.
63. Hieracium bauhini Bess.
64. Doronicum hungaricum (Saal) Rehb.
65. Lapsana communis L.
66. Mycelis muralis (L.) Dum.
67. Asparagus tenuifolius Lam.
68. Lilium martagon L.
69. Polygonatum latifolium (Jacq.) Desf.
70. Ornithogalum pyramidale L.
71. Ruscus aculeatus L.
72. Scilla bifolia L.
73. Arum orientale M.B.
74. Brachypodium sylvaticum (Huds.) Pal.- Blauv.
75. Hordelymus europaeus (L.) Jenssen
76. Melica uniflora Rerz.
77. Carex sylvatica Huds.
78. Carex pilosa Scop.
79. Himantoglossum hircinum (L.) Sprun.
80. Listera ovata (L.) R. Br.
81. Neottia nidus-avis (L.) L.C. Rich.
82. Platanthera bifolia (L.) L.C. Rich.
83. Epipactis latifolia (L.) All.
II. Rezervatia cu Paeonia peregrina
1. Ophioglossum vulgatum L.
3. Salix cinerea L.
4. Carpinus betulus L.
5. Quercus pedunculiflora C. Koch.
6. Quercus pubescens Wild.
7. Quercus robur L.
8. Ulmus foliacea Gilib.
9. Ligustrum vulgare L.
10. Cornus sanguinea L.
11. Evonimus verrucosa Scop.
12. Tilia tomentosa Moench.
13. Tilia platyphylos Scop.
14. Rosa canina L.
15. Rosa galica L.
16. Acer campestre L.
17. Acer tataricum L.
18. Acer platanoides L.
19. Pirus piraster (L.) Med.
20. Crataegus monogyna Jacq.
21. Holosteum umbellatum L.
22. Moehringia trinervia L. Clairv.
23. Anemone nemorosa L.
24. Anemone ranunculoides L.
25. Ficaria verna Huds.
26. Helleborus odorus W. et K.
27. Isopyrum thalictroides L.
28. Paeonia peregrina Mill. Var. romanica (Brandza) A. Nyar
29. Thalictrum aquilegifolium L.
30. Asarum europaeum L.
31. Hypericum hirsutum L.
32. Corydalis cava (L.) Schweigg. Et Koerte
33. Corydalis cava ssp. marchalliana (Pall.) Hay.
34. Corydalis solida (L.) Sn.
35. Alliaria petiolata (Bieb.) Cavara et Grande
36. Dentaria bulbiflora L.
37. Turritis glabra L.
38. Sedum maximum (L.) Hoffm.
39. Geum urbanum L.
40. Fragaria vesca L.
41. Potentilla alba L.
42. Astragalus glycyphyllos L.
43. Lathyrus niger (L.) Bernh.
44. Lathyrus vernus (L.) Bernh.
45. Geranium phaeum L.
46. Geranium robertianum L.
47. Euphorbia amygdaloides L.
48. Mercurialis perennis L.
49. Viola canina L.
50. Viola elatior Fries.
51. Viola sylvestris Lam.
52. Aegopodium podagraria L.
53. Chaerophyllum temulum L.
54. Vinca herbacea W. et K.
55. Asperula taurina L.
56. Lithospermum purpureo-coeruleum L.
57. Pulmonaria officinalis L.
58. Galeobdolon luteum Huds.
59. Galeopsis speciosa Mill.
60. Stachys sylvatica L.
61. Lathraea squamaria L.
62. Physalis alkekengi L.
63. Solanum nigrum L.
64. Scrophularia nodosa L.
65. Veronica chamaedrys L.
66. Valeriana officinalis L.
67. Campanula persicifolia L.
68. Campanula ranunculoides L.
69. Doronicum hungaricum (Saal) Rehb.
70. Lapsana communis L.
71. Mycelis muralis (L.) Dum.
72. Allium ursinum L.
73. Asparagus tenuifolius Lam.
74. Convallaria majalis L.
75. Gagea lutea (L.) Ker. - Gawl.
76. Lilium martagon L.
77. Polygonatum latifolium (Jacq.) Desf.
78. Ruscus aculeatus L.
79. Scilla bifolia L.
80. Galanthus nivalis L.
81. Leucojum aestivum L.
82. Croccus flavus Weston ssp. flavus
83. Iris variegata L.
84. Arum orientale M.B.
85. Brachypodium sylvaticum (Huds.) Pal. - Beauv.
86. Hordelymus europaeus (L.) Jenssen
87. Millium effusum L.
88. Carex sylvatica Huds.
89. Listera ovata (L.) R. Br.
90. Orchis purpurea Huds.
91. Platanthera bifolia (L.) L.C. Rich
92. Epipactis latifolia (L.) All.
SPECII DE NEVERTEBRATE INVENTARIATE IN BALTA COMANA
Filum Plathelminthes
Clasa Turbellaria
Ordinul Rhabdocoela
Prorhynchus stagnalis M. Schultze
Mesostoma ehrenbergi Falke
Mesostoma lingua Abildgard
Gyratrix hermafroditus Ehrenberg
Dalyellia viridis G. Shaw
Typhloplana viridata Abildgard
Ordinul Triclada
Dugesia lugubris O. Schmidt
Polycelis felina
Polycelis nigra O.F. Muller
Dendrocoelum lacteum O.F. Muller
Filum Nemathelminthes
Clasa Rotifera
Proalides tentaculatus de Beauchamp
Epiphanes clavulata Ehrenberg
Epiphanes senta O.F. Muller
Platyas patulus O.F. Muller
Brachionus quadridentatus Hermann
Rotaria elongata Weber
Epiphanes branchionus Ehrenberg
Filum Annelida
Clasa Oligochaeta
Stylaria lacustris
Clasa Hirudinea
Eropobdella nigricolis (Brandes, 1900)
Glossiphonia complanata (Linnaeus, 1758)
Piscicola geometra (Linnaeus, 1758)
Hirudo medicinalis (Linnaeus, 1758)
Haemopis sanguisuga (Linnaeus, 1758)
Clasa Insecta
Ordinul Diptera
Familia Chironomidae
Subfamilia Tanypodidae
Tnypus villipennis Kieff
Subfamilia Chironominae
Tribul Chironomini
Chironomus plumosus (Linnaeus, 1758)
IHTIOFAUNA CARACTERISTICA CURSULUI INFERIOR AL NEAJLOVULUI SI ZONEI BALTII COMANA
Desi caracterizata printr-o mare varietate de biotopi si biocenoze, zona Baltii Comana si segmentul inferior al raului Neajlov adapostesc un numar relativ redus de specii de pesti dintre care cele mai multe sunt caracterizate prin populatii slab reprezentate ca numar de indivizi. Cauzele probabile ale existentei unui numar redus de specii ar putea fi gradul crescut de toxicitate a apei raului Neajlov, constatata cu ocazia prelevarii de probe in vederea determinarii calitatii apei, cand s-au constatat concentratii crescute pentru anumiti ioni (nitriti, amoniu, fosfati), o importanta fluctuatie a unor parametri fizici acvatici, ca urmare a adancimii scazute a apei in lacurile formate de catre raul Neajlov, dezvoltarea in masa a organismelor planctonice (mai ales alge si bacterii) care cauzeaza fenomene de hipoxie in perioada de vara, intensificarea proceselor de descompunere a materiei organice care amplifica scaderea concentratiei oxigenului dizolvat si, nu in ultimul ran, fenomenul pradarii.
Avand in vedere existenta unor populatii mici de pesti, in cea mai mare parte fara importanta economica se poate aprecia ca aceasta nu prezinta o atractie deosebita din punct de vedere a pescuitului sportiv, cel putin in situatia actuala.
Dintre speciile de pesti capturate in zona aflata in studiu mentionam:
Supraclasa Pisces
Clasa Osteichthyes Goodrych 1909
Ordinul Clupeiformes Berg 1940
Subordinul Esocoidei Berg 1936
Familia Umbridae Gunter 1866
Genul Umbra Walbaum 1792
Umbra krameri Walbaum 1792
Familia Esocidae Gunter 1866
Genul Esox Linnaeus 1758
Esox lucius Linnaeus 1758
Ordinul Cypriniformes Goodrich 1909
Subordinul Cyprinoidei Berg 1940
Familia Cyprinidae Jordan st Evermann 1896
Genul Rutilus Rafinesque 1820
Subgenul Rutilus Rafinesque 1820
Rutilus rutilus carpathorossicus Vladycov 1930
Genul Leuciscus Cuvier 1817
Subgenul Squalius Bonaparte 1841
Leuciscus cephalus cephalus (Linnaeus) 1758
Leuciscus borysthenicus ssp. celensis (?)
Genul Scardinius Bonaparte 1837
Scardinius erythrophthalmus erythrophthalmus (Linnaeus) 1758
Genul Alburnus Heckel 1843
Alburnus alburnus (Linnaeus) 1758
Genul Rhodeus Agassiz 1776
Rhodeus sericeus amarus (Bloch) 1782
Genul Gobio Cuvier 1817
Gobio gobio Valenciennes 1844
Genul PseudorasboraBleeker 1860
Pseudorasbora parva (Schlegel) 1842
Genul Cyprinus Linnaeus 1847
Cyprinus carpio Linnaeus 1758
Genul Carassius Jarocki 1822
Carassius auratus gibelio (Bloch) 1783
Familia Cobitidae
Genul Misgurnus Lacepede 1803
Misgurnus fossilis Linnaeus 1758
Genul Cobitis Linnaeus 1758
Cobitis taenia taenia Linnaeus 1758
Cobitis danubialis
Ordinul Perciformes Bertin et Arambourg 1958
Familia Centrarchidae Regan 1913
Genul Lepomis Rafinesque 1819
Lepomis gibossus (Linnaeus ) 1758
Familia Percidae Jordan et Evermann 1896
Genul Perca Linnaeus 1758
Perca fluviatilis Linnaeus 1758
Genul Gymnocephalus
Gymnocephalus cernuus
Subordinul Gobioidei Berg 1911
Familia Gobiidae Regan 1911
Genul Neogobius Iljin 1927
Neogobius fluviatilis fluviatilis Pallas 1811
Dintre speciile prezentate, cateva au mare importanta pentru ihtiofauna Romaniei:
1. Leuciscus borysthenicus formeaza populatii stabile in zone restranse din Delta Dunarii. La Comana s-a identificat prezenta acestui gen, reprezentat insa printr-o forma asemenatoare, descrisa ca L. borysthenicus ssp. celensis, care formeaza o populatie stabila in paraul Valea Gurbanului, tributar raului Neajlov.
2. Misgurnus fossilis a cunoscut un important regres populational in Romania urmare a aridizarii generale a climei si a disparitiei habitatuluicaracteristic, dar si prin activitatile de amenajare a bazinelor hidrografice in scopul cresterii suprafetelor de teren destinate agriculturii.
3. O alta specie ale carei efective populationale s-au redus este si Esox lucius. O situatie similara a fost constatata pentru Umbra krameri.
In ultimele decenii in zona baltii Comana si-au facut aparitia si unele dintre speciile de ciprinide asiatice introduse in cultura in tara noastra, mentionand in acest sens Ctenopharyngodon idella (Valenciennes), Hypophthalmichthys molitrix (Valenciennes) si Aristichthys nobilis Richardson.
CLASA AMPHIBIA(ZONA BALTII COMANA SI PADUREA COMANA)
Ordinul Urodela
Familia Salamandridae
Triturus vulgaris vulgaris (Linnaeus) 1758
Ordinul Anura
Familia Discoglossidae
Bombina bombina (Linnaeus) 1761
Familia Pelobatidae
Pelobates fuscus fuscus (Laurentus) 1768
Bufo viridis viridis (Laurentus) 1768
Hyla arborea arborea (Linnaeus) 1758
Rana ridibunda ridibunda (Pallas) 1771
Rana esculenta (Linnaeus) 1758
Rana dalmatina (Bonaparte) 1839
CLASA REPTILIA
Ordinul Chelonia
Familia Emydidae
Emys orbicularis (Linnaeus) 1758
Ordinul Squamata
Subordinul Sauria
Familia Scincidae
Ablepharus kitaibelii fitzingeri (Mertens) 1952
Familia Lacertidae
Lacerta agilis chersonensis (Andrzejowski) 1832
Lacerta viridis viridis (Laurenti) 1768
Anguis fragilis colchicus (Nordmann) 1840
Subordinul Ophidia
Familia Colubridae
Coronella austriaca austriaca (Laurenti) 1768
Natrix natrix natrix (Linnaeus) 1758
Natrix tessellata tessellata (Laurenti) 1768
CLASA AVES
Ordinul Podicipediformes
Familia Podicipedidae
Genul Podiceps
Podiceps cristatus Linnaeus
Podiceps nigricolis Brehm
Podiceps ruficolis Pallas
Ordinul Pelecaniformes
Familia Phallacrocoracidae
Genul Phalacrocorax Brisson
Phalacrocorax carbo sinensis Show et Nodder
Ordinul Ciconiformes
Familia Ardeidae
Genul Ardea Linnaeus
Ardea cinerea Linnaeus
Ardea purpurea Linnaeus
Genul Ardeola Boie
Ardeola rallloides Scopoli
Genul Egretta Forster
Egretta alba Linnaeus
Genul Nycticorax Forster
Nycticorax nycticorax Linnaeus
Genul Botaurus Stephens
Botaurus stellaris Linnaeus
Familia Ciconidae
Genul Ciconia Linnaeus
Ciconia nigra Linnaeus
Ordinul Anseriformes
Familia Anatidae
Genul Cignus Bechstein
Cygnus olor Gmelin
Genul Anser Brisson
Anser anser Linnaeus
Anser albifrons Linnaeus
Genul Anas Linnaeus
Anas platyrhynchos
Anas strepera Linnaeus
Anas querquedula Linnaeus
Ordinul Charadriiformes
Subordinul Lari
Familia Laridae
Genul Larus Linnaeus
Larus argentatus Pontoppidan
Larus ridibundus Linnaeus
Familia Sternidae
Genul Chlidonias Rafinesque
Chlidonias niger Linnaeus
Genul Sterna Linnaeus
Sterna hirundo Linnaeus
Subordinul Charadrii
Familia Charadriidae
Genul Vanellus Brison
Vanellus vanellus Brisson
Familia Recurvirostridae
Genul Gallinago Brisson
Gallinago gallinago Linnaeus
Familia Scolopacidae
Genul Scolopax Linnaeus
Scolopax rusticola Linnaeus
Ordinul Gruiformes
Familia Rallidae
Genul Fulica Linnaeus
Fulica atra Linnaeus
Genul Gallinula Brison
Gallinula chloropus Linnaeus
Genul Crex Brechstein
Crex crex Linnaeus
Familia Rallidae
Genul Rallus Linnaeus
Ralus aquaticus Linnaeus
Ordinul Galliformes
Familia Phaseanidae
Genul Perdix Brisson
Perdix perdix Linnaeus
Genul Coturnix Bonnaterre
Coturnix coturnix Linnaeus
Genul Phaseanus Linnaeus
Phaseanus colchicus Linnaeus
Ordinul Columbiformes
Familia Columbidae
Genul ColumbaLinnaeus
Columba palumbus Linnaeus
Genul Streptopelia Bonaparte
Streptopelia decaocto Frivaldzsky
Ordinul Falconiformes
Familia Accipitridae
Genul Accipiter Brisson
Accipiter gentilis Linnaeus
Accipiter nissus Linnaeus
Genul Buteo Lacepede
Buteo buteo Linnaeus
Genul Circus Lacepede
Circus aeruginosus Linnaeus
Familia Falconidae
Genul Falco Linnaeus
Falco peregrinus Tunstall
Falco subbuteo Linnaeus
Falco tinnunculus Linnaeus
Falco vespertinus Linnaeus
Ordinul Strigiformes
Familia Strigidae
Genul Asio Brisson
Asio otus Linnaeus
Genul Strix Linnaeus
Strix aluco Linnaeus
Genul Athene Boie
Athene noctua Scopoli
Genul Otus Pennaut
Otus scops Linnaeus
Ordinul Cuculiformes
Familia Cuculidae
Genul Cuculus Linnaeus
Cuculus canorus Linnaeus
Ordinul Piciformes
Familia Picidae
Genul Dendrocopos Koch
Dendrocopos major Linnaeus
Dendrocopos medius linnaeus
Dendrocopos minor Linnaeus
Dendrocopos syriacus Hemprich/Ehrenberg
Genul Picus Linnaeus
Picus viridis Linnaeus
Picus canus Gmelin
Genul Dryocopus Linnaeus
Dryocopus martius Linnaeus
Genul Picoides Lacepede
Jynx torquilla Linnaeus
Ordinul Caprimulgiformes
Familia Caprimulgidae
Genul Caprimulgus Linnaeus
Caprimulgus europaeus Linnaeus
Ordinul Apodiformes
Familia Apodidae
Genul Apus Scopoli
Apus apus Linnaeus
Ordinul Coraciiformes
Familia Coracidae
Genul Coracias Linnaeus
Coracias garrulus Linnaeus
Genul Merops Brisson
Merops apiaster Linnaeus
Familia Alcedinidae
Genul Alcedo Linnaeus
Alcedo atthis Linnaeus
Familia Upupidae
Genul Upupa
Upupa epops linnaeus
Ordinul Passeriformes
Familia Muscicapidae
Genul Muscicapa Brisson
Muscicapa striata Pallas
Familia Sylviidae
Genul Sylvia Scopoli
Sylvia communis Latham
Sylvia nisoria Bechstein
Genul Acrocephalus Naumann
Acrocephalus arundinaceus Linnaeus
Acrocephalus scirpaceus Hermann
Acrocephalus schoenobaenus Linnaeus
Acrocephalus palustris Bechstein
Genul locustella Kaup
Locustella luscinioides Wolf
Locustella fluviatilis Wolf
Genul Hippolais Baldenstein
Hippolais icterina Vieillot
Hippolais pallida Hemprida/Ehrenberg
Genul Phyloscopus Boie
Phylloscopus collybita Veillot
Genul Turdus Linnaeus
Turdus merula Linnaeus
Turdus philomelos Brehm
Genul Oenanthe Vieillot
Oenanthe oenanthe Linnaeus
Genul Saxicola Bechstein
Saxicola rubetra Linnaeus
Luscinia megarhynchos Brehm
Luscinia luscinia Linnaeus
Genul Phoenicurus Forster
Phoenicurus phoenicurus Linnaeus
Genul Erithacus Cuvier
Erithacus rubecula Linnaeus
Familia Laniidae
Genul Lanius Linnaeus
Lanius colurio Linnaeus
Lanius excubitor Linnaeus
Familia Paridae
Genul Parus Linnaeus
Parus major Linnaeus
Parus caeruleus Linnaeus
Familia Aegithalidae
Genul Aegithalos Hermann
Aegithalos caudatus Linnaeus
Familia Hirundinidae
Genul Hirundo Linnaeus
Hirundo rustica Linnaeus
Genul Riparia Forster
Riparia riparia riparia Linnaeus
Genul Delichon Horsfield/Moore
Delichon urbica urbica Linnaeus
Familia Sittidae
Genul Sitta Linnaeus
Sitta europaea caesia Wolf
Familia Fringillidae
Genul Fringilla Linnaeus
Fringilla coelebs coelebs Linnaeus
Genul Carduelis Brisson
Carduelis carduelis Linnaeus
Carduelis chloris chloris Linnaeus
Genul Coccothraustes Brisson
Coccothraustes coccothraustes Linnaeus
Genul Emberiza Linnaeus
Emberiza citrinella citrinella Linaeus
Genul Miliaria
Miliaria calandra calandra Linnaeus
Familia Passerinae
Genul Passer Brisson
Paser domesticus Linnaeus
Paser domesticus Linnaeus
Paser montanus montanus Linnaeus
Familia Alaudidae
Genul Alauda Linnaeus
Alauda arvensis cantarella Bonaparte
Genul Galerida Boie
Galerida cristata cristata Linnaeus
Familia Motacillidae
Genul Motacilla Linnaeus
Motacilla alba alba Linnaeus
Motacilla flava Linnaeus
Familia Sturnidae
Genul Sturnus Linnaeus
Sturnus vulgaris Linnaeus
Familia Oriolidae
Genul Oriolus Linnaeus
Orilus oriolus Linnaeus
Familia Corvidae
Genul Garrulus Brisson
Garrulus glandarius glandarius Linnaeus
Genul Pica Brisson
Pica pica pica Linnaeus
Genul Corvus Linnaeus
Corvus frugilegus Linnaeus
Corvus corone cornix Linnaeus
Corvus monedula Linnaeus
CARACTERISTICI ALE AVIFAUNEI IN ZONA BALTII COMANA
Complexitatea conditiilor stationale oferite de varietatea formelor de relief, hidrografice, edafice etc. confera intregii zone un aspect mozaicat, populat de o mare varietate de specii apartinand avifaunei. Functie de aceste caracteristici si transformarile permanente ale ecosistemului complex care caracterizeaza regiunea Comana, si popularea acestor zone cu specii va fi determinata de exigentele indivizilor asupra habitatului dar si conditiile de mediu in permanenta transformare. Astfel, teritoriile nou formate prin depunere de aluviuni si colmatare, instabile, nu vor fi populate de pasari pentru cuibarit insa ele constituie locuri excelente pentru hranire, ceea ce explica inventarierea unui numar relativ mare de specii de pasari, cu populatii reduse numeric, dar putine cuibaritoare.
Stabilizarea aluviunilor prin fixarea unor asociatii vegetale constituite predominant din genurile Typha si Phragmites au creat conditii de adapost si reproducere pentru un numar insemnat de specii caracteristice zonelor umede: rate, starci, egrete, nagati, lisite, gainuse dar si numeroase specii de paseriforme. In locurile mai inalte ale cuvetei lacustre, asociatiile vegetale de stuf si papura cuprind si salcii pitice, locuri ideale de cuibarit pentru coloniile de starci, apartinand mai multor genuri existente in zona. Pe solul ramas la suprafata apei ca urmare a lucrarilor de regularizare a cursului Neajlovului s-a fixat chiar vegetatie arborescenta, reprezentata prin salcam si diferite specii de arbori caracteristici zonelor umede, in special plopi si salcii. Albia minora a raului Neajlov, cu maluri lutoase, situate la aproximativ 1,5 metri deasupra nivelului apei pe o anumita distana constituie, de asemenea, locul predilect de hranire pentru unele specii limicole.
Salciile si plopii plantate in vederea stabilizarii malurilor si stoparii inaintarii stufului constituie locuri de cuibarit, dar si puncte de observatie pentru numeroase specii de pasari de prada. Acelasi rol important il detine liziera padurii din localitatea Comana care formeaza limita sudica a baltii Comana pe o distanta de aproximativ 6 km. Varsta apreciabila a unor arbori, mai ales stejari, ca si portul acestora ofera conditii optime de cuibarit pentru numeroase specii de pasari ale caror afinitati biologice corespund caracteristicilor de interferenta dintre ecosistemele lacustre si forestiere, dar si specii proprii fiecaruia dintre cele doua habitate.
Dintre speciile care se concentreaza in zona de liziera a padurii se mentioneaza pradatorii diurni, mai multe specii de ciocanitori, iar dintre paseriforme, mai ales genul Parus.
Luciul de apa, desi in suprafata mica constituie zone de concentrare pentru mai multe specii de rate.
Malul dezgolit din stanga tehnica a Neajlovului, cu zone de teren cultivate agricol si putina vegetatie lemnoasa subarbustiva, constituie loc de refugiu pentru unele pasari apartinand unor specii de Phaseanidae, dar si pentru mierle, sturzi, sfrancioci.
In timp, componenta biocenozelor s-a modificat considerabil, mai ales in ultimele doua decenii, sub presiunea transformarii biotopurilor caracteristice, a reducerii drastice a suprafetei luciului de apa, astfel specii care anterior erau foarte bine reprezentate ca marime a populatiilor fiind in prezent caracterizate printr-un numar redus de indivizi, cu prezenta pe o perioada foarte scurta de timp in zona.
OBSERVATII ASUPRA FENOLOGIEI PASARILOR DIN COMANA
In complexul Comana se intalnesc patru categorii fenologice principale: specii oaspeti de vara (54% din numarul total de specii); specii oaspeti de iarna (12% din numarul total de specii); specii sedentare (28% din totalul speciilor) si specii de pasaj (6% din totalul de specii prezente in zona). (fig. 1).
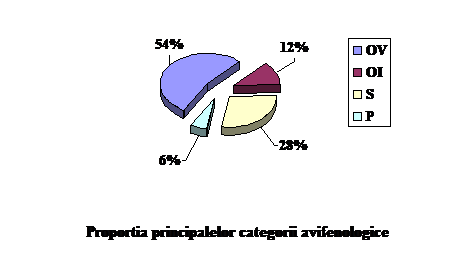
Analizandu-se provenienta geografica a oaspetilor de iarna si a oaspetilor de vara se constata ca speciile din prima categorie apartin tipurilor de fauna arctic si siberian, iar cele din a doua categorie apartin tipurilor de fauna european, mongol si mediteranean. Aceste specii apartin in mare la doua tinuturi geografice, unul nordic si unul sudic.
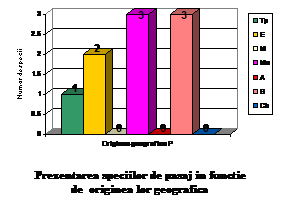
Total
Ponderea %
Tp
33
21.56
E
75
49
M
10
6.53
Mo
16
10.45
A
3
1.96
S
14
9.15
Ch
2
1.3
total
153
100
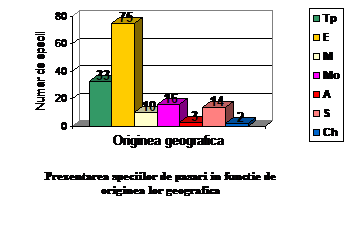
O parte din aceste specii cuibaresc departe in nord si sosesc in Comana fie ca forme de iarna, fie ca forme de pasaj. Aceste specii sunt "oaspeti de iarna" si au origine nordica. Intalnim 19 astfel de specii (12% din totalul speciilor observate).
Cealalta parte din specii, vin primavara din sud pentru a se reproduce, vizitand Comana numai in sezonul cald, pentru ca apoi odata cu inceperea racirii timpului sa se intoarca pentru iernat in sud. Aceste specii "oaspeti de vara" sunt de origine sudica. Intalnim 82 astfel de specii (54% din totalul speciilor observate).
Speciile de pasaj, includ speciile care apar numai in timpul migratiei, primavara spre locurile de reproducere din nord iar toamna spre locurile de iernare din sud. Se intalnesc 9 astfel de pecii (6% din totalul speciilor observate).
Speciile sedentare intrunesc un numar de 43 de specii (28% din totalul speciilor observate) a caror prezenta este semnalata in tot cursul anului.
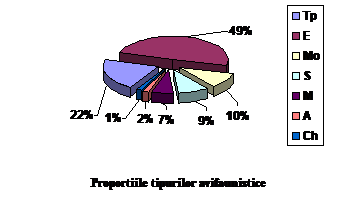
Analizand tipurile geografice la care apartin speciile din cele patru categorii avifenologice se constata urmatoarele aspecte:
OASPETII DE IARNA
Speciile oaspeti de iarna sunt forme arctice, siberiene, europene si transpalearctice. Astfel intalnim: 3 specii de origine arctica (15.7 % din totalul speciilor oaspeti de iarna); 10 specii de origine siberiana (52.63 % din totalul speciilor oaspeti de iarna); 3 specii de origine transpalearctica (15.7 % din totalul speciilor oaspeti de iarna) si 3 specii de origine europeana (15.7 % din totalul speciilor oaspeti de iarna)
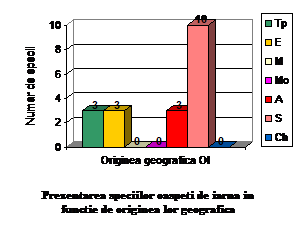
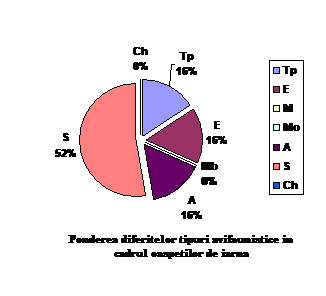
Orig.geog.
OI
Ponderea %
Tp
3
15.78
E
3
15.78
M
0
0
Mo
0
0
A
3
15.78
S
10
52.63
Ch
0
0
total
19
100
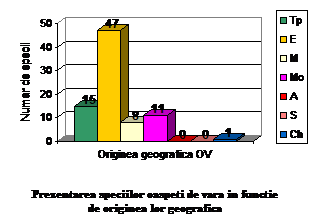
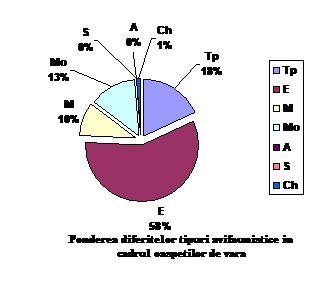
Orig. geog.
OV
Ponderea %
Tp
15
18.29
E
47
57.31
M
8
9.75
Mo
11
13.41
A
0
0
S
0
0
Ch
1
1.21
total
82
100
OASPETII DE VARA
Speciile oaspeti de vara apartin indeosebi formelor europene , transpalearctice, mongole si mediteraneene. Astfel intalnim: 47 specii de origine europeana (57.31 % din totalul speciilor oaspeti de vara); 15 specii de origine transpalearctica (18.29 %din totalul speciilor oaspeti de vara); 11 specii de origine mongola (13.41 % din totalul speciilor oaspeti de vara); 8 specii de origine mediteraneana (9.75 % din totalul speciilor oaspeti de vara) si o specie de origine chineza ( 1.21 % din totalul speciilor oaspeti de vara).
SPECIILE DE PASAJ
Speciile de pasaj cuprind urmatoarele categorii: 3 specii de origine siberiana (33.33 % din totalul speciilor de pasaj); 3 specii de origine mongola (33.33 % din totalul speciilor de pasaj); 2 specii de origine europeana (22.22 % din totalul speciilor de pasaj); o specie de origine transpalearctica (11.11 % din totalul speciilor de pasaj);
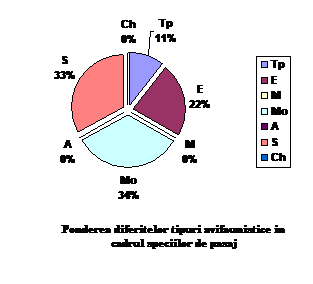
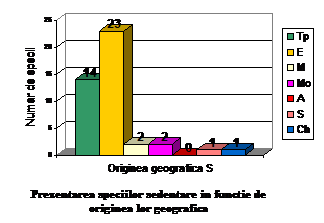
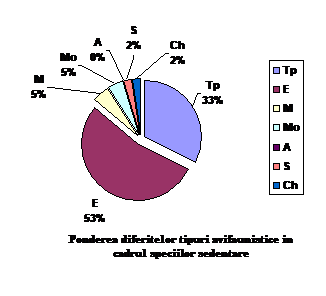
Orig geog.
S
Ponderea %
Tp
14
32.55
E
23
53.48
M
2
4.65
Mo
2
4.65
A
0
0
S
1
2.32
Ch
1
2.32
total
43
100
Orig. geog.
P
Ponderea %
Tp
1
11.11
E
2
22.22
M
0
0
Mo
3
33.33
A
0
0
S
3
33.33
Ch
0
0
total
9
100
SPECIILE SEDENTARE
Speciile sedentare cuprind urmatoarele categorii: 23 specii de origine europeana ( 53.48 % din totalul speciilor sedentare); 14 specii de origine transpalearctica ( 32.55 % din totalul speciilor sedentare); 2 specii de origine mediteraneana (4.65 % din totalul speciilor sedentare); 2 specii de origine mongola (4.65 % din totalul speciilor sedentare); o specie de origine siberiana ( 2.32 % din totalul speciilor sedentare) si o specie de origine chineza ( 2.32 % din totalul speciilor sedentare).
CARACTERIZAREA HABITATELOR SI BIOTOPURILOR
1. Habitatul acvatic
1.a. Biotopul lacustru este reprezentat de suprafata de aproximativ 6 ha a doua lacuri de mica adancime ( sub 1 metru adancime in perioada de vara) si a canalelor permanente care fac legatura intre aceste ochiuri de apa sau intre ele si raul Neajlov. Suprafata luciului de apa creste insa in perioadele cu precipitatii abundente, functie de debitul raului Neajlov. Aceste lacuri constituie teritoriile de hranire pentru unele dintre speciile prezente (rate, lisite, gainuse de balta), iar vegetatia densa din jurul lor ofera conditii bune de cuibarit pentru acestea. Viteza de circulatie a apei este scazuta mai ales in perioada de vara cand legaturile cu riul sunt partial intrerupte, iar improspatarea apei este dificil de realizat. In aceasta situatie se produce dezvoltarea exploziva a organismelor planctonice (fito- si zooplanctonice), dar in mod special a algelor si bacteriilor.
1.b. Raul Neajlov se caracterizeaza printr-un coeficient mare de sinuozitate in zona baltii, formand meandre si numeroase canale derivate, de mica adancime (cu apa permanenta sau temporar irigate). Cursul Neajlovului este caracterizat printr-o viteza de scurgere a apei destul de mare, cu o adancime care variaza de la cativa zeci de centimetri in zona din aval pana la 2 - 3 metri in unele zone situate la coturile albiei minore si o latime medie cuprinsa intre 4 si 6 metri in zona situata amonte fata de balta Comana, ajungand la 10 - 12 metri aval fata de aceasta. Cursul raului este flancat pe cea mai mare distanta de stuf si in unele locuri, cu apa mai putin adanca si viteza de scurgere redusa, de papura, pentru ca in zona din aval, pe o distanta de aproximativ 500 m, malurile sa nu fie acoperite de vegetatie inalta.
2. Stufarisurile (zonele acoperite de stuf si papura) inundate permanent cu apa, de mica adancime, in unele locuri existand si salcii pitice ofera conditii ideale de cuibarit pentru unele specii coloniale, de exemplu pentru starci. Fiind greu accesibil pentru oameni si pradatori, linistit, acest habitat ofera si conditii bune de hranire pentru majoritatea speciilor care il populeaza. Functie de adancimea apei, raportul dintre cele doua genuri de plante (Typha si Phrogmites) se modifica, conferind un anumit grad de varietate a habitatului.
Habitat amfibiu
3. Liziera padurii, habitat aflat la interferenta ecosistemului acvatic cu cel terestru cuprinde zone inundate temporar de ape. Pe marginea padurii Comana, in proximitatea baltii, exista stejari seculari, de inaltimi si diametre mari care constituie locuri posibile de cuibarit si puncte de observatie pentru numeroase specii, mai ales pradatori.
Sub scoarta sau in tesutul lemnos isi sapa galerii larvele de insecte, acesti arbori constituind locuri de hranire pentru numeroase specii de pasari insectivore (mai ales Picidae). In zonele de limita a padurii unde s-au efectuat lucrari de exploatare a masei lemnoase a aparut un bogat lastaris, constituind zone de concentrare a unor specii de paseriforme.
4. Plantatiile de salcii si plopi de pe malul stang al Neajlovului, inundate temporar de apa, cu numeroase izvoare care mentin o umiditate ridicata in tot cursul anului asigura dezvoltarea unei vegetatii erbacee bogate, asigurand totodata locuri de cuibarire si hranire pentru numeroase pasari.
5. Pasunile, aparute urmare a scaderii nivelului apei si regularizarii cursului, amplasate in regiunea inferioara si superioara a baltii Comana se afla deasupra nivelului obisnuit al apei, fiind inundate numai in timpul viiturilor. Pasunile se caracterizeaza in general prin vegetatie erbacee, dar in locurile mai joase, temporar inundate de apa se dezvolta papura si uneori stuf. Avand suprafete relativ reduse si fiind pasunate nu ofera conditii deosebite de cuibarire sau hranire pentru pasari.
Habitat terestru
Padurea Comana
6. Padurea compacta este caracterizata in zona situata in imediata apropiere a baltii prin arbori varstnici, mai ales tei si frasini, dar si stejari de dimensiuni apreciabile, cu scorburi si ramuri uscate.
7. Malul stang al baltii Comana, acoperit in unele locuri cu vegetatie arbustiva si subarbustiva, de talie mica, ofera conditii de adapost, liniste si cuibarire pentru unele specii de pasari. Caracterizate prin pante mari in anumite zone, straturile superficiale de sol au fost partial erodate. Avand expozitie insorita asigura locuri de cuibarit pentru speciile care depun pontele in galerii sapate in malurile argiloase.
Raportat la intregul habitat acest biotop este insa nesemnificativ ca suprafata si importanta ca zona de cuibarit.
Biotopurile mentionate anterior au existenta permanenta sau temporara. Importanta lor pentru speciile de pasari prezente difera functie de suprafata lor (care variaza in mod direct cu nivelul apei raului Neajlov) si avantajele pe care le ofera pentru hranire si reproducere. O importanta deosebita se pot mentiona trei dintre biotopuri descrise anterior si anume: stufarisurile, liziera padurii si luciul de apa al locurilor si canalelor de legatura dintre acestea.
CARACTERIZAREA ORNITOLOGICA A HABITATELOR SI BIOTOPURILOR
Incadrarea speciilor de pasari intr-un anumit tip de habitat sau biotop specific se realizeaza pe baza afinitatilor biologice ale indivizilor fata de caracteristicile generale ale acestora, ca teritorii de cuibarit, hranire sau adapostire.
Daca pentru numeroase specii de pasari teritoriul de hranire este altul decat cel de cuibarit, dar in cadrul aceluiasi habitat, pentru altele cele doua teritorii (biotopuri) se suprapun in totalitate (paseriforme).
O alta categorie de specii o formeaza cele care intr-o anumita perioada din ciclul biologic folosesc mai multe biotipuri, din habitate diferite (de exemplu unele specii de pradatori de zi care cuibaresc in zona forestiera, dar teritoriul de hranire se suprapune cel putin partial peste zona baltii).
In cazul speciilor care utilizeaza mai multe biotopuri ele vor fi descrise in acela care le caracterizeaza prin densitatea cea mai mare de indivizi, avand in acelasi timp o prezenta constanta, ceea ce corespunde perioadei de cuibarire.
Habitatele speciilor de vara
Habitatul acvatic
Suprafetele ochiurilor de apa si canalelor care fac legatura intre acestea nu caracterizeaza prezenta permanenta a pasarilor, ele fiind populate decat temporar pentru ca nu ofera conditii de cuibarire. Aceste biotipuri ofera insa conditii optime de hranire pentru speciile de rate, gainuse de balta, lisite. Benzile de papura (mai ales in zonele unde exista vegetatie uscata, din anii precedenti, la suprafata apei) ofera conditii ideale de cuibarit pentru speciile mentionate.
1. Speciile caracteristice lacurilor si canalelor deschise in interiorul stufului:
- Podiceps cristatus
- Podicep griseigena
- Podiceps nigricollis
- Anas strepera
- Anas querquedulla
- Larus ridibundus
- Sterna hirundo
- Fulica atra
2. Speciile caracteristice stufarisurilor compacte si palcurilor de salcii pitice
Ardea cinerea
Ardea purpurea
Ardea ralloides
Egreta alba
Nycticorax nycticorax
Aceste specii cuibaresc de obicei in colonii mixte, in interiorul suprafetelor compacte de stuf in care apar salcii pitice sau stufarisuri culcate pe sol sau la suprafata apei, in zone putin deranjate prin activitati antropice sau de pradatori specifici (dintre mamifere). Teritoriile de hranire sunt localizate in mediul acvatic pentru starcul cenusiu si starcul rosu, pentru ca celelalte specii mentionate in acest biotop sa isi gaseasca hrana in mediul amfibiotic (zona de mal si alte teritorii irigate temporar), hrana fiind preponderent acvatica.
Speciile caracteristice habitatului acvatic sunt in majoritate migratoare ; chiar putinele specii sedentare (cele care isi au asigurata hrana din resursele acvatice) parasesc balta in iernile geroase, cand apa ingheata complet sau se concentreaza in ochiurile de apa ramase neinghetate de pe raul Neajlov (rata mare, pescarusul argintiu, pescarusul razator).
Habitatul amfibiu
Desi populat cu indivizi apartinand la numeroase specii care caracterizeaza alte habitate este bine reprezentat faunistic, data fiind abundenta si varietatea hranei din acest habitat.
Dintre speciile care populeaza plantatiile de plopi si salcii pot fi citate:
Muscicapa striata
Sylvia atricapilla
Luscinia luscinia
Phoenicurus phoenicurus
Parus major
Parus coeruleus
Fringilla coelebs
Corvus corone cornix
Diversitatea mare de specii localizate in acest biotop este explicata prin varietatea locurilor de cuibarit (in scorburi, in coroanele salciilor, in vegetatia abundenta de la sol) dar si prin siguranta locurilor de cuibarit si diversitatea si abundenta hranei.
Un alt biotop caracteristic habitatului amfibiu este reprezentat de stufarisurile vechi si papurisurile bogate in maracinisuri care acopera, in situatia actuala, cea mai mare parte din suprafata baltii. Caracteristice acestui biotop sunt speciile:
Bataurus stellaris
Ixobrychus minutus
Gallinula chloropus
Circus aeruginosus
Acrocephalus palustris
Emberiza schoeniclus
Cuiburile sunt amplasate in vegetatia densa, deasupra apei sau pe sol. Malurile lutoase ale raului Neajlov, in regiunea lipsita de stuf si papura, aval fata de balta este caracterizata prin putine specii de pasari.
Motocilla alba
Motacilla flava
Larus ridibundus
Acest biotop constituie locul de hranire pentru specii apartinand Familiei Ardeidae.
Speciile prezentate cuibaresc rar in aceasta zona, depunand ouale pe sol sau in palcurile de vegetatie din zonele mlastinoase.
Pasunile, biotop de suprafata relativ mica raportat la intregul ecosistem al baltii este ocupat de un numar mic de specii in zona baltii Comana, una dintre cauzele care conduc la diminuarea populatiilor fiind pasunatul dar si uscarea temporara a acestor suprafete. Dintre speciile prezente se mentioneaza Rallus aquaticus, Matacilla alba si Motacilla flava, precum si specii apartinand genului Porzana.
Majoritatea speciilor de pasari observate in acest tip de habitat sunt migratoare sau sunt forme de pasaj. In timpul iernilor, speciile sedentare care populeaza pasunile umede din apropierea baltii fie ca se vor concentra in habitatul acvatic (rata mare), fie ca se vor deplasa spre plantatiile de salcii, plopi, dar cele mai multe vor fi atrase de conditiile climatice mai uniforme din interiorul padurii.
Habitatul terestru
De o mare complexitate a biotopilor in zona baltii Comana, habitatul terestru cuprinde un numar insemnat de specii, dintre care numeroase sedentare, dar si multe forme care apar in aceasta zona in perioada de iarna. Dintre biotopii caracteristici acestui tip de habitat este mentionat tipul ,,pasune uscata", care caracterizeaza zona de limita a baltii Comana din partea stanga tehnica a raului Neajlov.
Speciile caracteristice acestui biotop sunt:
Phasianus calchicus mangolicus
Perdix perdix
Coturnix coturnix
Alauda arvensis
Galerida cristata
Anthus campestris
Teritoriile de cuibarit ale acestor specii sunt reprezentate de terenurile agricole cu vegetatie erbacee si maracinisurile localizate pe pantele abrupte ale malurilor mai inalte. Ariile de hranire sunt localizate in proximitatea cuiburilor. Hrana este mixta (natura animala si vegetala).
Biotopul malurilor abrupte, de importanta redusa ca suprafata, specii gazduite si marime a populatiilor, reprezinta locul de cuibarire pentru Merops apiaster si Riparia riparia. Hrana acestor specii este de provenienta mixta, fiind preluata de la distante mai mici sau mai mari fata de zona de cuibarit.
Habitatul forestier, care se intinde in lungul baltii Comana, pe o distanta de aproximativ 6 km, formand limita sudica a acesteia este un tip de padure caracteristic zonei de campie joasa, constituita dintr-un mare numar de specii de arbori, predominand insa teiul, frasinul si stejarul. Caracteristica unitatii de productie din apropierea baltii Comana este varsta inaintata a arborilor (stejari si frasini seculari), care compun liziera padurii oferind locuri de cuibarire, observatie si hranire pentru multe specii. Dintre speciile prezente in acest biotop, cu frecventa mare apar:
Phasianus colchicus
Columba palumbus
Accipiter gentilis
Accipiter nisus
Buteo buteo
Buteo lagopus
Falco tinnunculus
Falco vespertinus
Asio otus
Dendrocopos major
Dendrocopos medius
Dendrocopos minor
Picus canus
Picus viridis
Caprimulgus europaeus
Upupa epops
Coracias garrulus
Sylvia communis
Sylvia curruca
Turdus merula
Turdus phylomelos
Luscinia megarhynchos
Erithacus rubecula
Lanius collurio
Parus palustris
Sitta europaea
Carduelis carduelis
Carduelis chloris
Coccothraustes coccothraustes
Paser montanus
Anthus trivialis
Sturnus vulgaris
Oriolus oriolus
Pica pica
Garrulus glandarius
Marea majoritate a pasarilor din acest habitat au un spectru larg de hranire, utilizand hrana atat de natura animala cat si vegetala, pe care o procura din habitate si biotopi diferiti.
Comparand cele trei tipuri de habitate pe baza componentei avifaunistice se constata o varietate foarte mare a speciilor pentru habitatul forestier, comparativ cu celelalte doua. Desi de suprafata foarte mare (mai mult de 1 000 hectare) balta Comana adaposteste un numar relativ restrans de specii raportat la alte ecosisteme acvatice similare. De asemenea, numarul mic de indivizi din cadrul populatiilor este o consecinta a transformarii generale a habitatului si modificarii conditiilor de viata pentru cele mai multe dintre speciile descrise inainte de anul 1990.
IDENTIFICAREA HABITATULUI FORESTIER AL PASARILOR
In cadrul padurii Comana s-au parcurs cateva trasee / transecte, indeosebi in aria protejata si in suprafetele cu arbori seculari (stejar, frasin etc.). O atentie deosebita s-a acordat lizierei, la intrepatrunderea ecosistemului forestier cu cel acvatic (balta Comana). Au fost notate toate speciile observate sau identificate dupa cantec/strigat si in cateva cazuri dupa cuiburi. Observatiile s-au realizat in cursul lunii iulie si la inceputul lunii august. Dupa centralizarea speciilor determinate s-au selectat cele care prezinta importanta pe plan local si global, respectiv acele pasari care se afla incluse in anexele unor conventii di directive internationale (Conventia de la Berna, Directiva Pasari a CE).
S-a constatat ca intreaga suprafata a padurii Comana prezinta importanta pentru pasarile clocitoare, toate habitatele si nisele existente fiind ocupate de pasari in epoca reproducerii.
S-au identificat patru (4) specii de importanta globala (Directiva Pasari a CE - anexa I): ciocanitoarea sura (Picus canus), ciocanitoarea pestrita mijlocie (Dendrocopos medius), muscarul gulerat (Ficedula albicollis) si sfranciocul rosietic (Lanius collurio).
Primele trei specii sunt dependente de existenta arborilor maturi/batrani, cu trunchiuri groase si a arborilor uscati sau debilitati pe care se hranesc. Pentru aceste specii este necesara o suprafata variabila de padure matura, din categoria codrului, asa cum sunt parcelele existente in rezervatii si in zona de liziera (la contactul dintre padure si balta). Aceste specii nu prezinta zone strict delimitate de clocit in cuprinsul acestor suprafete de padure batrana, perechile mutandu-se dintrr-un loc in altul functie de diversi factori. Asadar, nu se poate vorbi de o fidelitate fata de locul de cuibarit de la un an la altul.
Sfranciocul rosietic (Lanius collurio) cuibareste in tufe de regula spinoase de la liziera padurii sau din poieni. In padurea Comana exista cateva enclave unde s-a exploatat padurea sau au fost poieni mai vechi, precum si zone restranse cu maces (Rosa canina) si porumbar (Prunus spinosa) la liziera. Se constata astfel ca viata acestei pasari este legata doar de prezenta zonelor deschise ale padurii Comana.
Recomandari pentru protectia zonelor importante de cuibarit
1. Conservarea arborilor de peste 80 - 100 ani, exceptandu-se de la taiere zone de padure care au in componenta arbori varstnici, mai ales in zona de contact dintre padure si balta Comana. Se vor conserva in primul rand arborii din speciile: stejar (Quercus robur), frasin (Fraxinus excelsior) si cires (Prunus avium);
2. Pastrarea arborilor uscati, rupti, scorburosi, debilitati in vederea conservarii siturilor de cuibarit si hrana din perimetrul protejat (se recomanda pastrarea tuturor arborilor debilitati sau in diferite stari de degradare de la liziera, la contactul intre padure si balta Comana, pe o fasie cu latimea de cel putin 40 metri). Prin aceasta masura se va evita disparitia unor specii de pasari rare din padurea Comana, printre care si cateva rapitoare de noapte (ordinul Strigiformes), posibil cuibaritoare in acest perimetru.
3. Efectuarea lucrarilor de ingrijire si exploatare forestiera in afara sezonului de cuibarit, cuprins intre 15 martie - 1 septembrie (masura este recomandata pentru unitatile de ameliorare cu arbori batrani);
4. Evitarea aplicarii tratamentelor chimice de orice fel impotriva agentilor patogeni in padurea Comana si pe suprafata baltii;
5. Conservarea vegetatiei arbustive din poieni, parchete exploatate si mai ales de la liziera padurii, la contactul cu balta Comana. Se va conserva indeosebi macesul (Rosa canina) si alte specii arbustive cu spini pentru protejarea locurilor de cuibarit ale speciei Lanius collurio - sfrancioc rosietic;
6. Aplicarea masurilor de restrictionare a turismului in padure (amplasarea de panouri avertizoare, bariere pentru limitarea accesului).
CLASA MAMMALIA
Ordinul Fissipedes
Suprafamilia Canoidea
Familia Canidae
Vulpes vulpes Linnaeus
Familia Mustelidae
Purorius putorius Linnaeus
Mustela nivalis Linnaeus
Martes martes Linnaeus
Meles meles Linnaeus
Lutra lutra Linnaeus
Suprafamilia Feloidea
Felis silvestris Schreber
Ordinul Artiodactyla
Subordinul Suiformes
Familia Suidae
Sus scrofa Kelm
Subordinul Ruminantia
Familia Cervidae
Capreolus capreolus Linnaeus
Cervus elaphus Linnaeus
Ordinul Insectivora
Familia Erinaceidae
Erinaceus concolor (Martin) 1838
Familia Talpidae
Talpa europaea (Linnaeus) 1758
Familia Sorecidae
Sorex araneus (Linnaeus) 1758
Neomys anomalus (Cabrera) 1907
Crocidura leucodon (Normann) 1780
Crocidura snaveolens (Pallas) 1811
Ordinul Lagomorpha
Familia Leporidae
Lepus europaeus Pallas
Ordinul Rodentia
Familia Sciuridae
Sciurus vulgaris Linnaeus
Familia Cricetidae
Cricetus cricetus Linnaeus
Familia Microtidae (Arvicolidae)
Pitymus subterraneus (De Selys - Longchamps) 1836
Ondatra zibetica (Linnaeus) 1766
Familia Muridae
Rattus norvegicus (Berkenhout) 1769
Mus musculus (Linnaeus) 1758
Mus spicilegus (Petenyi) 1882
Apodemus agrarius (pallas) 1778
Apodemus sylvaticus (Linnaeus) 1758
Mycromis minutus (Pallas) 1771
Familia Myoxidae
Dryomis nitedula (Pallas) 1779
Glis glis (Linnaeus) 1758
Ordinul Chiroptera
Subordinul Microchiroptera
Familia Rinolophidae
Rinolophus ferrumequinum Schreber
Familia Vespertilionidae
Myotis myotis Borkhausen
CONCLUZII GENERALE ASUPRA BIODIVERSITATII ZONELOR CERCETATE
Biodiversitatea floristica si faunistica din regiunea de campie a fost in general putin cercetata. Din acest motiv foarte multi taxoni, rari si amenintati cu disparitia au fost semnalati in ultima vreme prin cercetari efectuate in zonele umede de campie din sudul tarii. Cercetari anterioare au fost executate de catre: prof. dr. Ion Cristureanu, lect. dr. Ionel Ciugulea, lect. dr. Niculae Craciun, lect. dr. Florin Aioanei, academician Petru Banarascu, dr. Teodor Nalbant, dr. bilog Maria Pauca, dr. biolog Petrescu Angela, biolog Hodor Calin, prof. dr. Doina Ivan.
Acad. Petru Banarascu si lect. dr. Niculae Craciun au descris o noua specie de clean(Leuciscus celensis) endemica la Comana. Probabil aceasta specie exista si in alte zone invecinate.
Prin prezentul contract de cercetare am stabilit prin cercetari de teren efectivele speciilor amenintate de plante si animale care se gasesc in zonele umede.
S-au efectuat cercetari in captivitate de reproducere a speciilor rare, cercetari de etologie si biologia reproducerii pentru a pune la punct tehnici de reproducere dirijata in vederea repopularii.
Zonele umede au fost relativ putin cercetate, motiv pentru care nu se cunostea inca biodiversiatatea lor reala si masura in care conditiile de habitat nu se schimba sub influenta antropica si a poluarii.
In decurs de trei ani echipa de la Facultatea de Biologie a propus numeroase zone umede ca rezervatii stiintifice in urma explorarilor de teren facute.
Cercetarile au evidentiat de asemenea efectivele speciilor rare si au permis realizarea unei baze de date fluente despre speciile rare, baza de date care serveste echipelor care vor realiza manegementul durabil si monitoringul economic al speciilor rare.
Tinerii cercetatori astazi doctoranzi si-au facut stagiul de cercetare de 4-10 ani in zona umeda Comana la inventarierea speciilor rare de plante si animale, cu precadere a speciilor de pesti, amfibieni, reptile si pasari.
Unii tineri s-au specializat in metode de laborator de investigatii la nivel microscopic (analiza parazitilor -mai ales ectoparazitii) si la nivel macroscopic: efectuarea injectarilor hormonale pt. stimularea reproducerii in captivitate.
In regiunea Comana exista un numar foarte mare de taxoni foarte rari si declarati amenintati la nivel european (Conventia de la Berna, programul "Natura 2000" a Consiliului Europei).
Rezultatele au fost valorificate prin publicarea acestora in reviste de specialitate din tara.